

Inhibition of EZH2 Reduces Aging-Related Decline in Interstitial Cells of Cajal of the Mouse Stomach
- PMID: 38969206
- PMCID: PMC11359770
- DOI: 10.1016/j.jcmgh.2024.101376
Inhibition of EZH2 Reduces Aging-Related Decline in Interstitial Cells of Cajal of the Mouse Stomach
Abstract
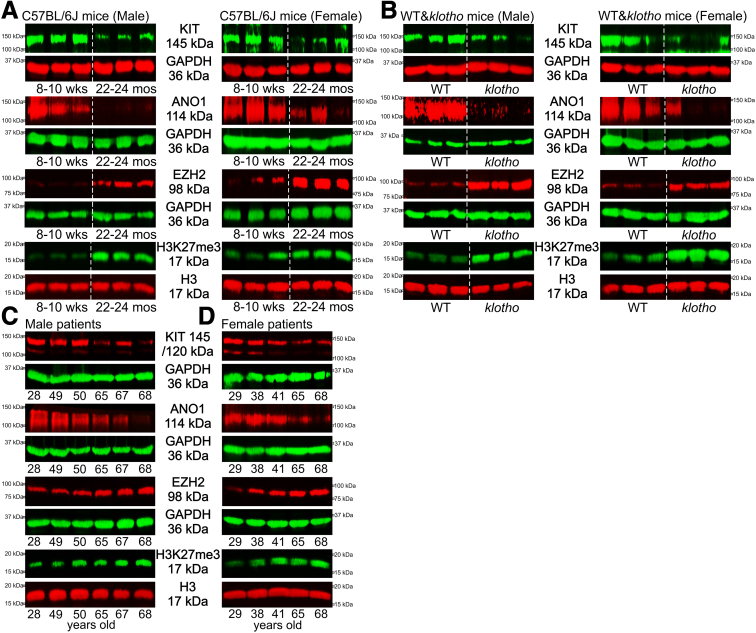
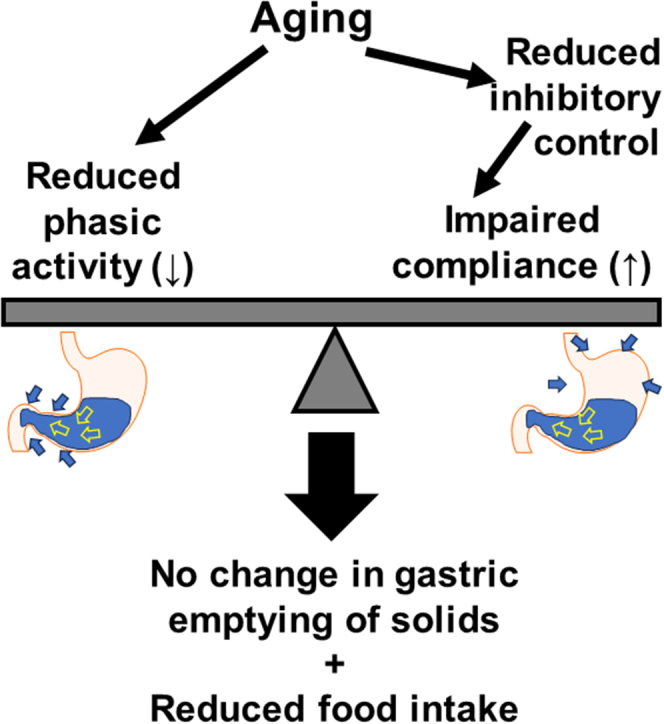
Background & aims: Restricted gastric motor functions contribute to aging-associated undernutrition, sarcopenia, and frailty. We previously identified a decline in interstitial cells of Cajal (ICC; gastrointestinal pacemaker and neuromodulator cells) and their stem cells (ICC-SC) as a key factor of gastric aging. Altered functionality of the histone methyltransferase enhancer of zeste homolog 2 (EZH2) is central to organismal aging. Here, we investigated the role of EZH2 in the aging-related loss of ICC/ICC-SC.
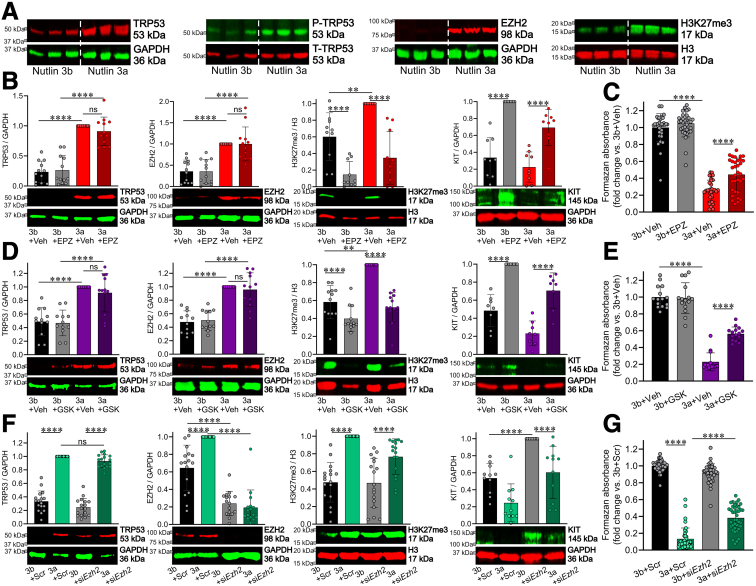
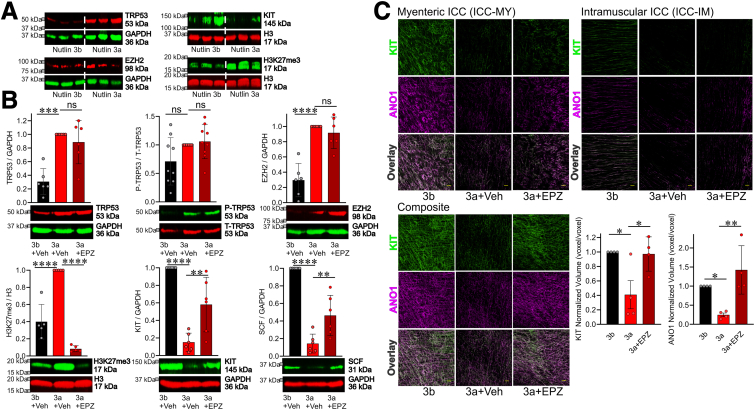
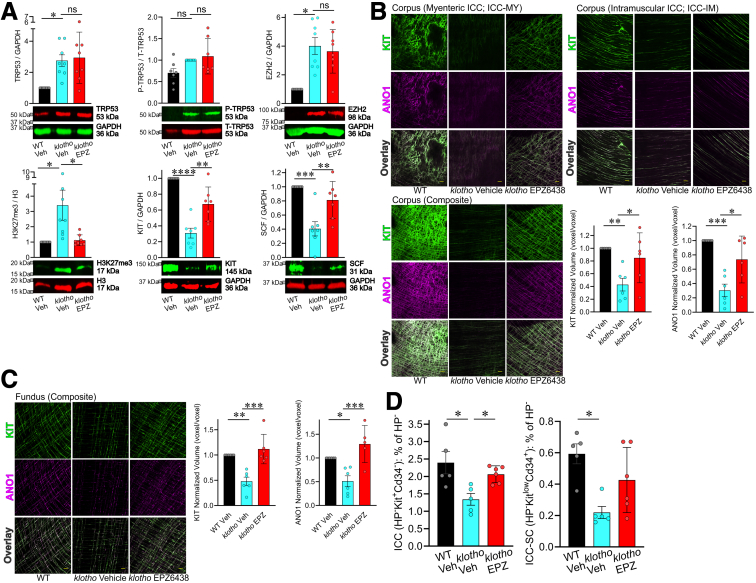
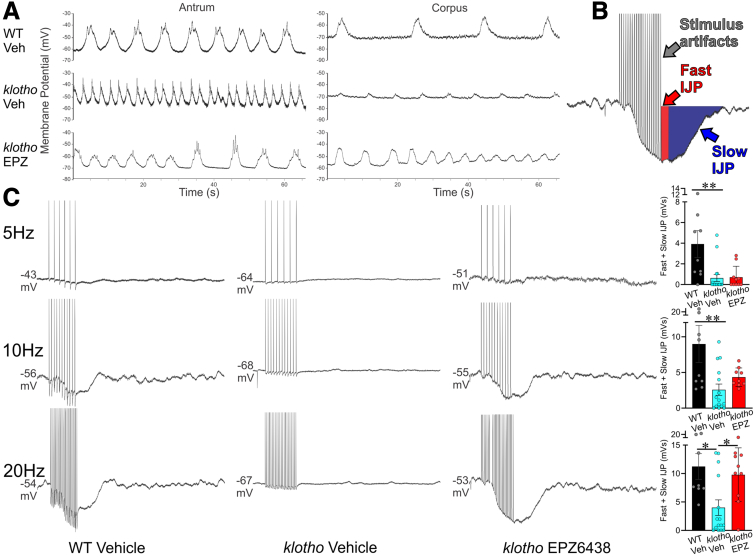
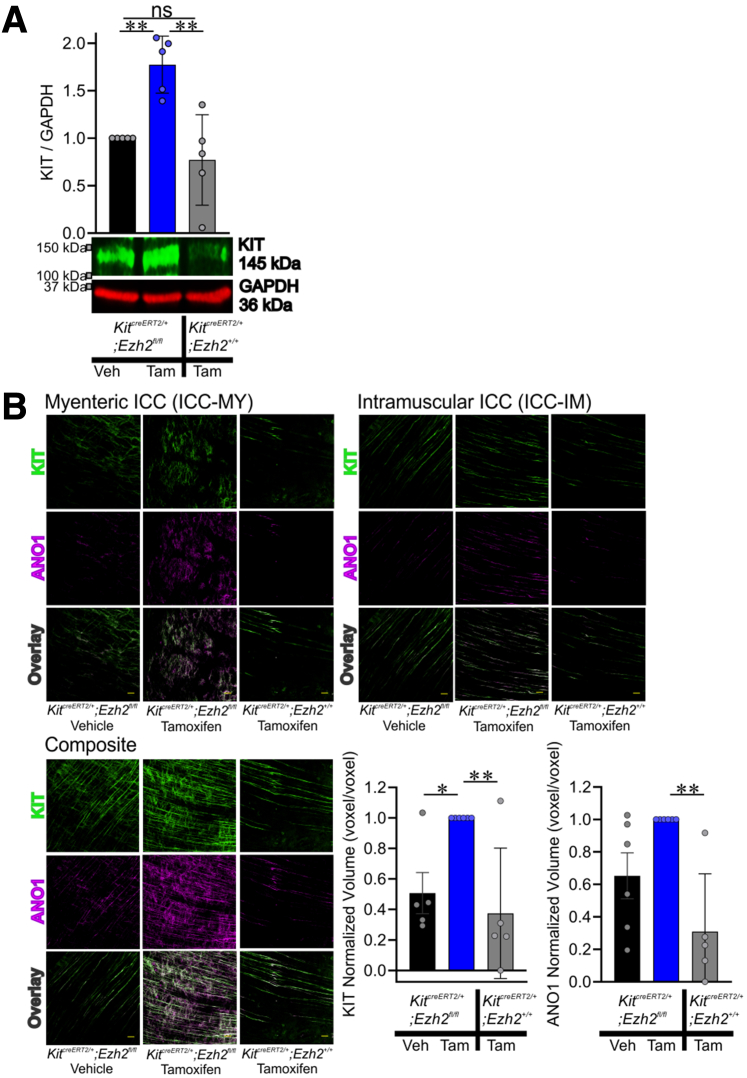
Methods: klotho mice, a model of accelerated aging, were treated with the most clinically advanced EZH2 inhibitor, EPZ6438 (tazemetostat; 160 mg/kg intraperitoneally twice a day for 3 weeks). Gastric ICC were analyzed by Western blotting and immunohistochemistry. ICC and ICC-SC were quantified by flow cytometry. Gastric slow wave activity was assessed by intracellular electrophysiology. Ezh2 was deactivated in ICC by treating KitcreERT2/+;Ezh2fl/fl mice with tamoxifen. TRP53, a key mediator of aging-related ICC loss, was induced with nutlin 3a in gastric muscle organotypic cultures and an ICC-SC line.
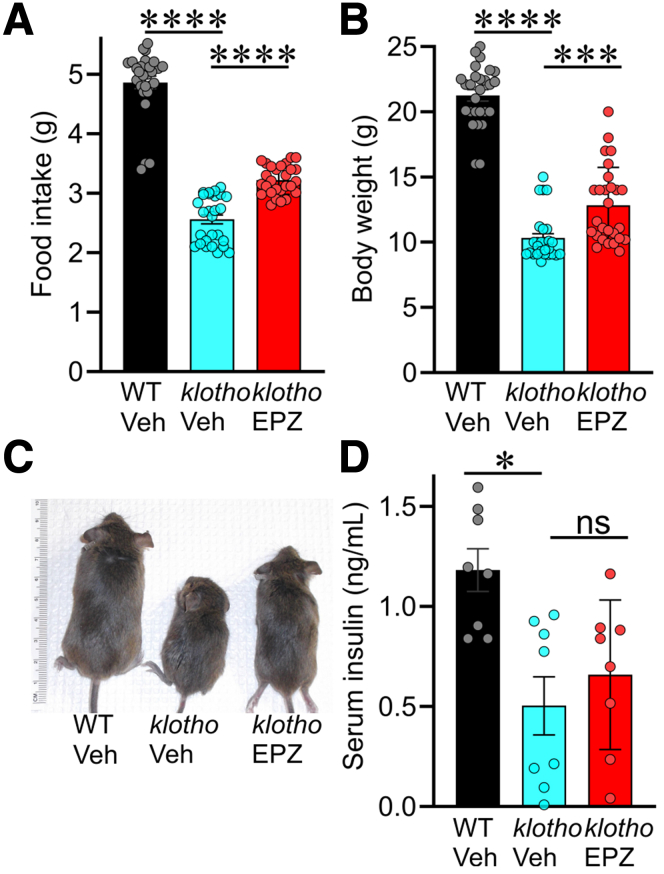
Results: In klotho mice, EPZ6438 treatment mitigated the decline in the ICC growth factor KIT ligand/stem cell factor and gastric ICC. EPZ6438 also improved gastric slow wave activity and mitigated the reduced food intake and impaired body weight gain characteristic of this strain. Conditional genomic deletion of Ezh2 in Kit-expressing cells also prevented ICC loss. In organotypic cultures and ICC-SC, EZH2 inhibition prevented the aging-like effects of TRP53 stabilization on ICC/ICC-SC.
Conclusions: Inhibition of EZH2 with EPZ6438 mitigates aging-related ICC/ICC-SC loss and gastric motor dysfunction, improving slow wave activity and food intake in klotho mice.
Keywords: Ezh2(fl/fl); Food Intake; Gastric Motility; Kit(creERT2/+); klotho.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
