

Early rhombic lip Protogenin+ve stem cells in a human-specific neurovascular niche initiate and maintain group 3 medulloblastoma
- PMID: 38971152
- PMCID: PMC11707800
- DOI: 10.1016/j.cell.2024.06.011
Early rhombic lip Protogenin+ve stem cells in a human-specific neurovascular niche initiate and maintain group 3 medulloblastoma
Abstract
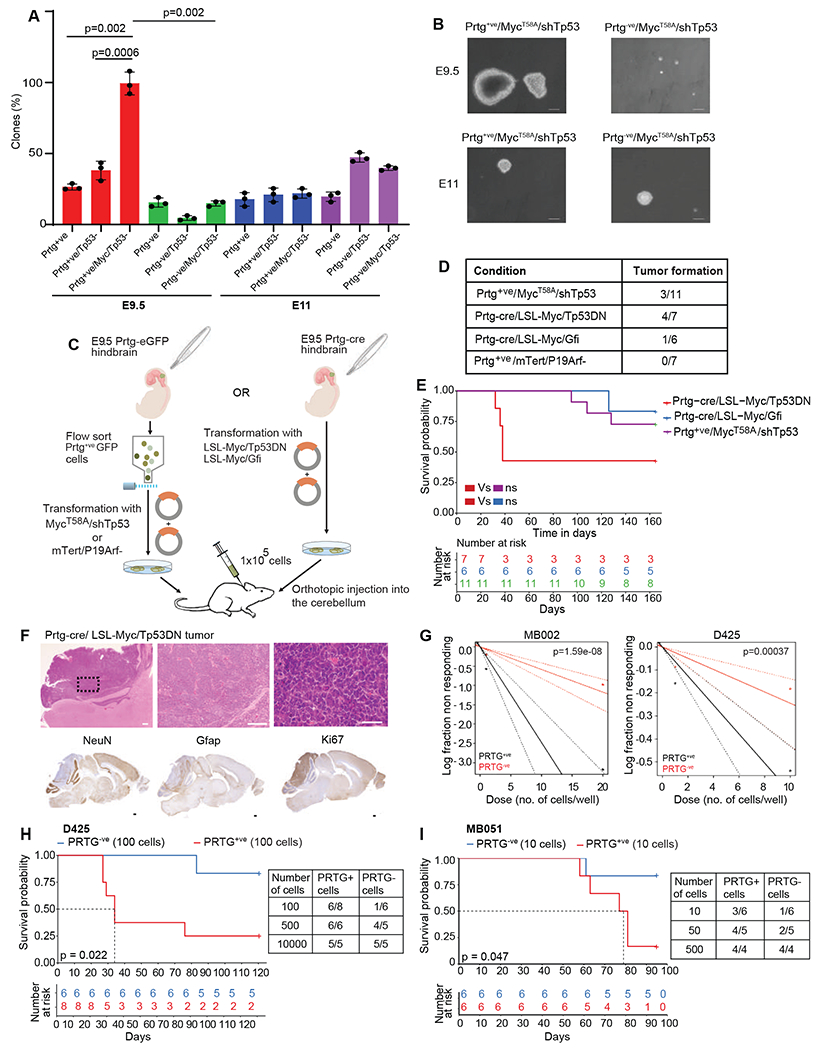
We identify a population of Protogenin-positive (PRTG+ve) MYChigh NESTINlow stem cells in the four-week-old human embryonic hindbrain that subsequently localizes to the ventricular zone of the rhombic lip (RLVZ). Oncogenic transformation of early Prtg+ve rhombic lip stem cells initiates group 3 medulloblastoma (Gr3-MB)-like tumors. PRTG+ve stem cells grow adjacent to a human-specific interposed vascular plexus in the RLVZ, a phenotype that is recapitulated in Gr3-MB but not in other types of medulloblastoma. Co-culture of Gr3-MB with endothelial cells promotes tumor stem cell growth, with the endothelial cells adopting an immature phenotype. Targeting the PRTGhigh compartment of Gr3-MB in vivo using either the diphtheria toxin system or chimeric antigen receptor T cells constitutes effective therapy. Human Gr3-MBs likely arise from early embryonic RLVZ PRTG+ve stem cells inhabiting a specific perivascular niche. Targeting the PRTGhigh compartment and/or the perivascular niche represents an approach to treat children with Gr3-MB.
Keywords: brain development; brain tumor immunotherapy; cancer genomics; group 3 medulloblastoma; perivascular niche; rhombic lip.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests. A patent related to this work was submitted by University of Pittsburgh (University Docket No. 05700/F&R ref. 48881-0028P01), US 63/275,326, “Molecules that bind to Protogenin polypeptides.”
Figures






References
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
