

Redox-dependent plasticity of oxMIF facilitates its interaction with CD74 and therapeutic antibodies
- PMID: 38972295
- PMCID: PMC11263951
- DOI: 10.1016/j.redox.2024.103264
Redox-dependent plasticity of oxMIF facilitates its interaction with CD74 and therapeutic antibodies
Abstract
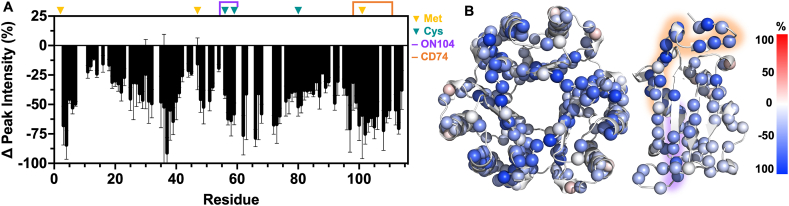
MIF is a ubiquitous protein involved in proinflammatory processes, which undergoes an oxidation-driven conformational change to oxidized (ox)MIF. We demonstrate that hypochlorous acid, produced by neutrophil-released myeloperoxidase (MPO) under inflammatory conditions, effectively oxidizes MIF into the oxMIF isoform, which is specifically recognized by the anti-oxMIF therapeutic antibody, ON104. NMR investigation of MIF oxidized by the MPO system revealed increased flexibility throughout the MIF structure, including at several catalytic and allosteric sites. Mass spectrometry of MPO-oxMIF revealed methionines as the primary site of oxidation, whereas Pro2 and Tyr99/100 remained almost unmodified. ELISA, SPR and cell-based assays demonstrated that structural changes caused by MPO-driven oxidation promoted binding of oxMIF to its receptor, CD74, which does not occur with native MIF. These data reveal the environment and modifications that facilitate interactions between MIF and its pro-inflammatory receptor, and a route for therapeutic intervention targeting the oxMIF isoform.
Keywords: CD74; Hypochlorous acid; Inflammation; MIF; Methionine; Myeloperoxidase; Neutrophils; ON104; oxMIF.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: At the time of the study, SS, MF, AS, IM, JM, GR and MT are employees of OncoOne R&D GmbH (Vienna, Austria), MT, and AS hold shares in OncoOne R&D GmbH, and MT and AS are inventors on a patent application related to the work described here.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
