

The antibodies against the A137R protein drive antibody-dependent enhancement of African swine fever virus infection in porcine alveolar macrophages
- PMID: 38973388
- PMCID: PMC11259084
- DOI: 10.1080/22221751.2024.2377599
The antibodies against the A137R protein drive antibody-dependent enhancement of African swine fever virus infection in porcine alveolar macrophages
Abstract
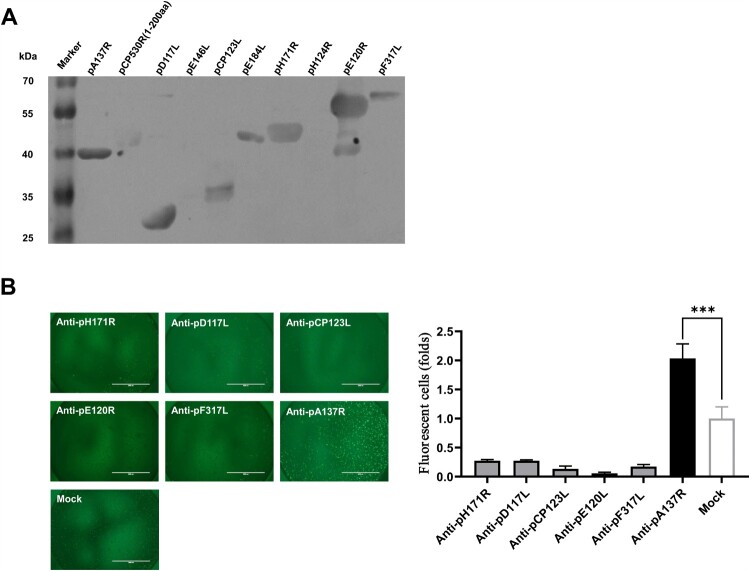
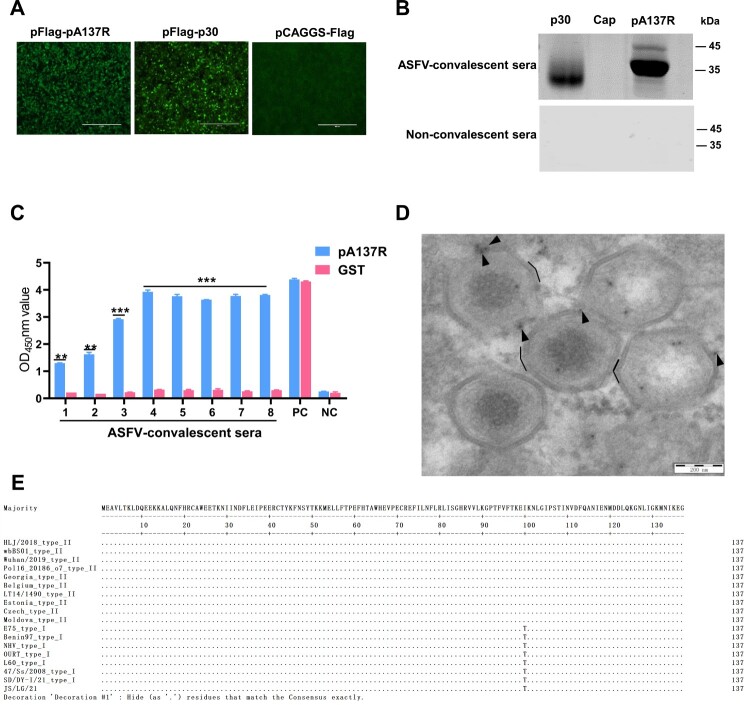
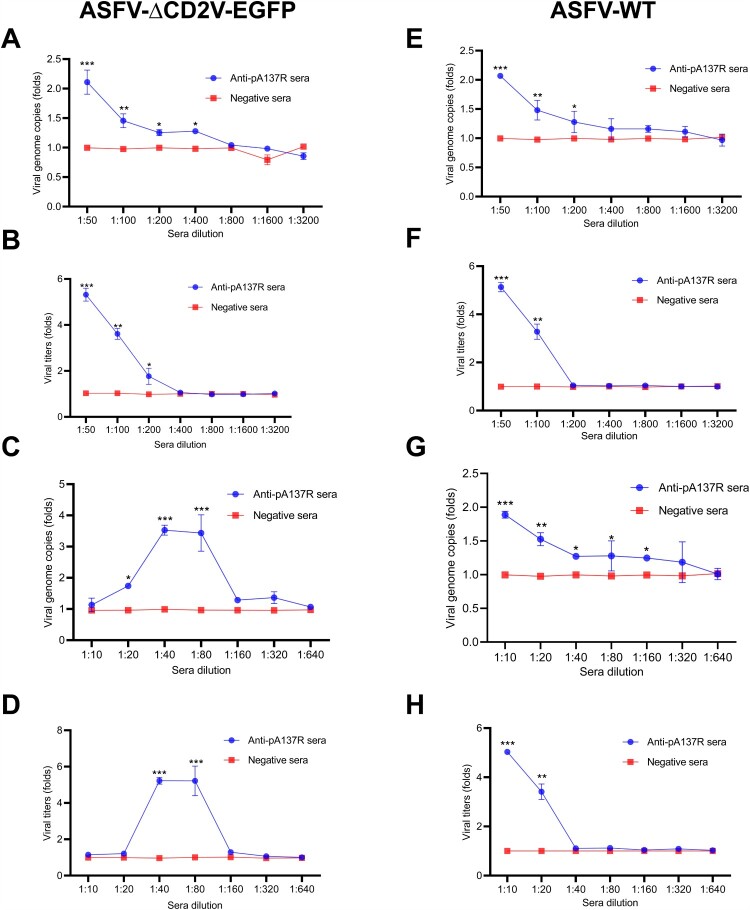
African swine fever virus (ASFV) is the causative agent of African swine fever (ASF), a highly contagious disease that can kill up to 100% of domestic pigs and wild boars. It has been shown that the pigs inoculated with some ASF vaccine candidates display more severe clinical signs and die earlier than do pigs not immunized. We hypothesize that antibody-dependent enhancement (ADE) of ASFV infection may be caused by the presence of some unidentified antibodies. In this study, we found that the ASFV-encoded structural protein A137R (pA137R) can be recognized by the anti-ASFV positive sera, indicating that the anti-pA137R antibodies are induced in the ASFV-infected pigs. Interestingly, our results demonstrated that the anti-pA137R antibodies produced in rabbits or pigs enhanced viral replication of different ASFV strains in primary porcine alveolar macrophages (PAMs), the target cells of ASFV. Mechanistic investigations revealed that anti-pA137R antibodies were able to promote the attachment of ASFV to PAMs and two types of Fc gamma receptors (FcγRs), FcγRII and FcγRIII, mediated the ADE of ASFV infection. Taken together, anti-pA137R antibodies are able to drive ASFV ADE in PAMs. These findings shed new light on the roles of anti-ASFV antibodies and have implications for the pathophysiology of the disease and the development of ASF vaccines.
Keywords: A137R protein; African swine fever virus; Fc gamma receptors; antibody-dependent enhancement; viral replication.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources