

CircHTT(2,3,4,5,6) - co-evolving with the HTT CAG-repeat tract - modulates Huntington's disease phenotypes
- PMID: 38974999
- PMCID: PMC11225910
- DOI: 10.1016/j.omtn.2024.102234
CircHTT(2,3,4,5,6) - co-evolving with the HTT CAG-repeat tract - modulates Huntington's disease phenotypes
Abstract
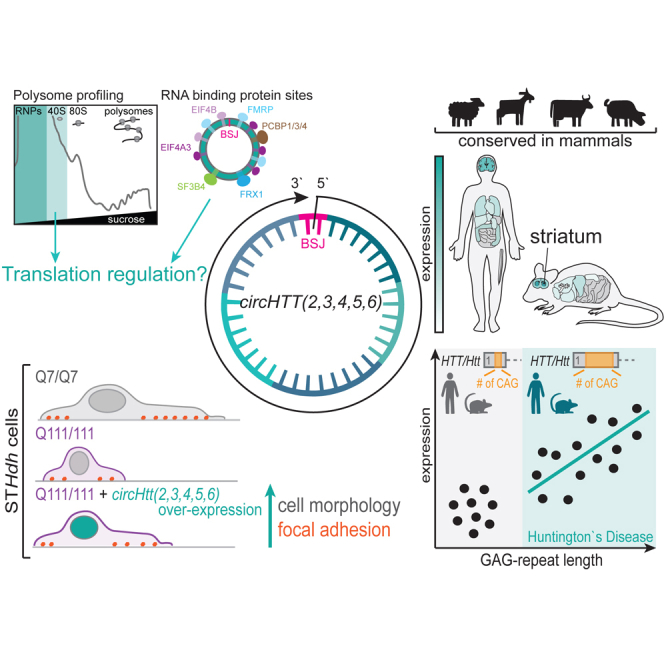
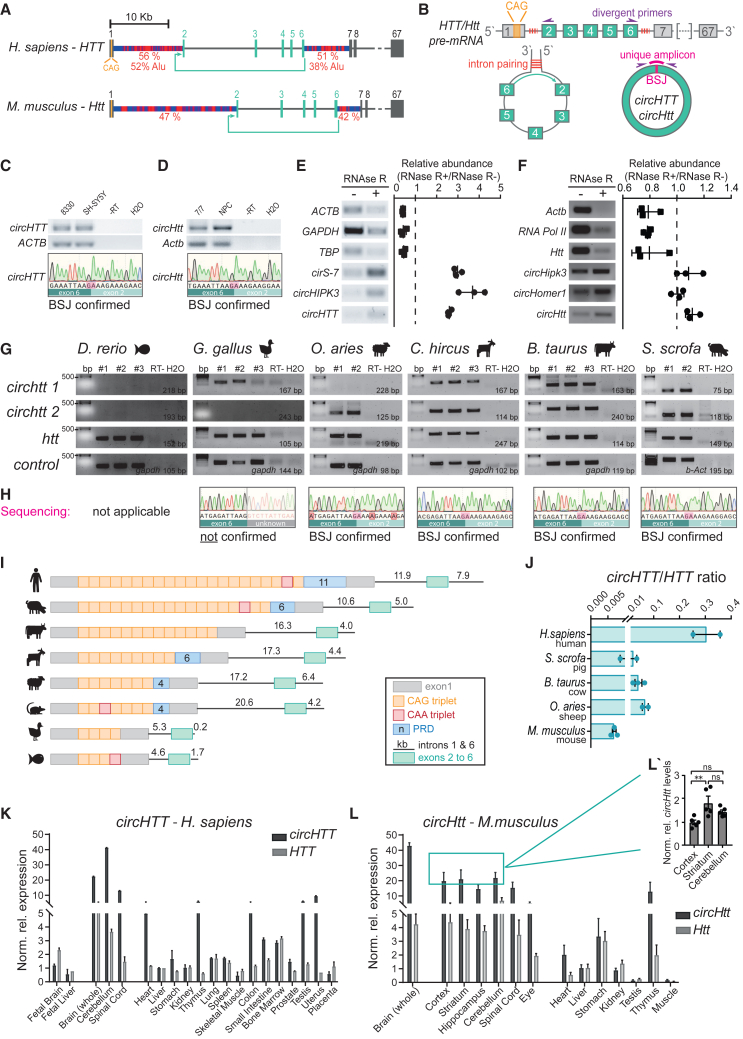
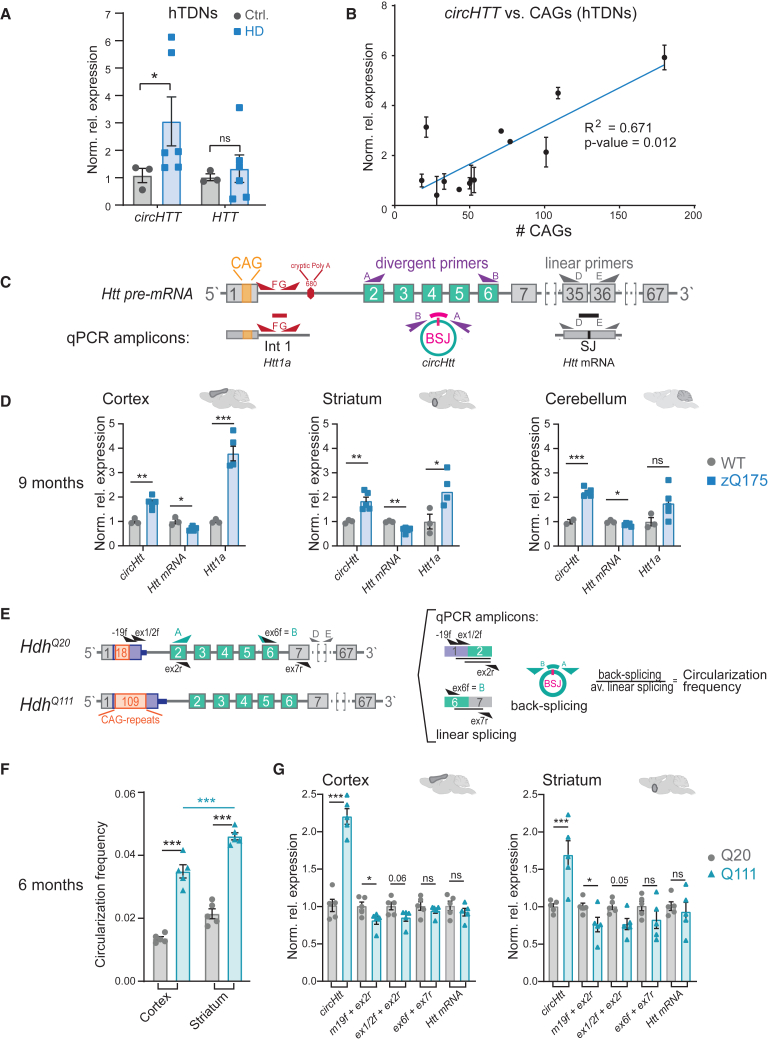
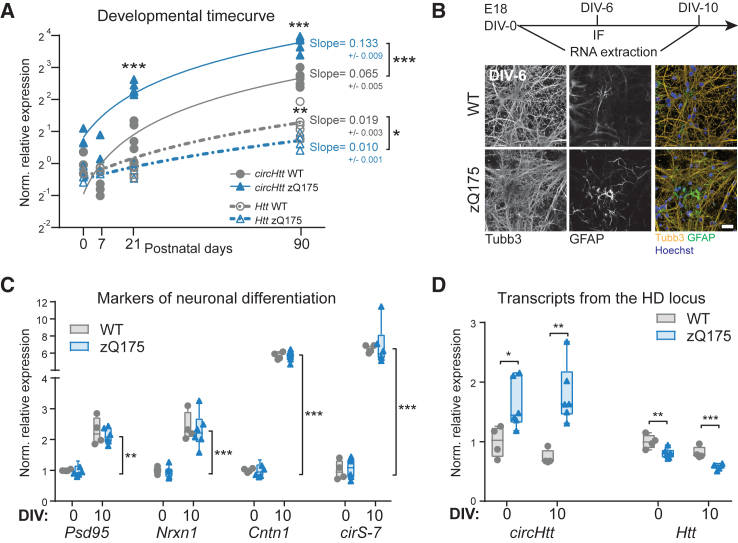
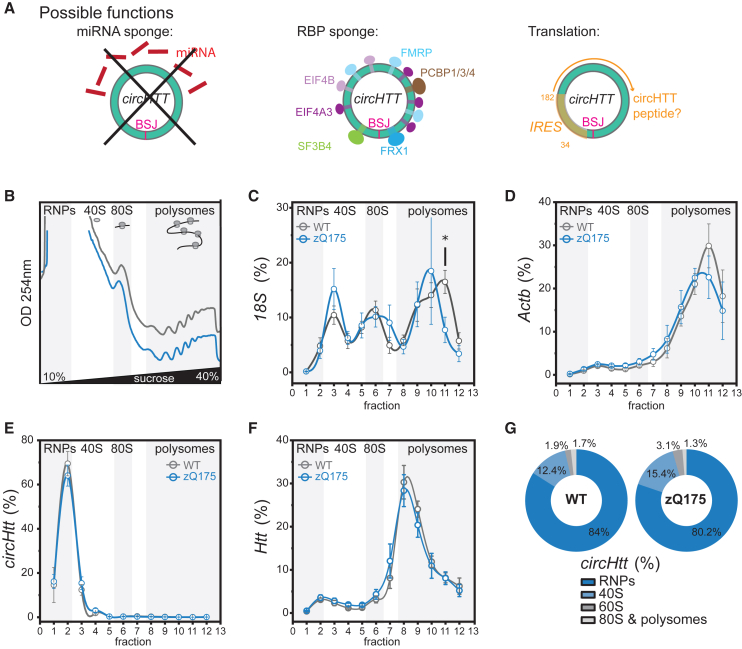
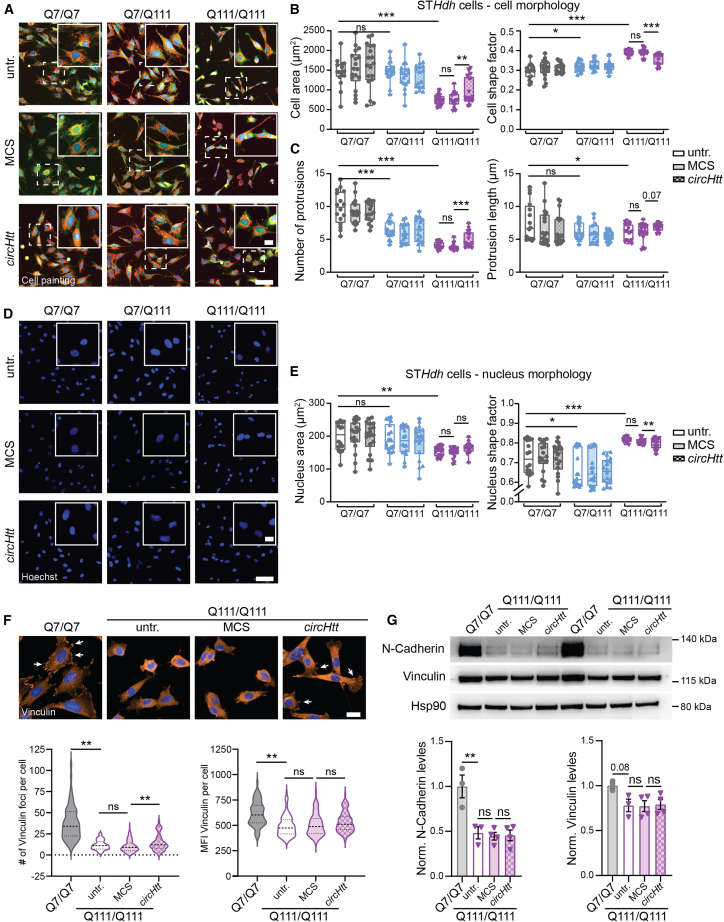
Circular RNA (circRNA) molecules have critical functions during brain development and in brain-related disorders. Here, we identified and validated a circRNA, circHTT(2,3,4,5,6), stemming from the Huntington's disease (HD) gene locus that is most abundant in the central nervous system (CNS). We uncovered its evolutionary conservation in diverse mammalian species, and a correlation between circHTT(2,3,4,5,6) levels and the length of the CAG-repeat tract in exon-1 of HTT in human and mouse HD model systems. The mouse orthologue, circHtt(2,3,4,5,6), is expressed during embryogenesis, increases during nervous system development, and is aberrantly upregulated in the presence of the expanded CAG tract. While an IRES-like motif was predicted in circH TT (2,3,4,5,6), the circRNA does not appear to be translated in adult mouse brain tissue. Nonetheless, a modest, but consistent fraction of circHtt(2,3,4,5,6) associates with the 40S ribosomal subunit, suggesting a possible role in the regulation of protein translation. Finally, circHtt(2,3,4,5,6) overexpression experiments in HD-relevant STHdh striatal cells revealed its ability to modulate CAG expansion-driven cellular defects in cell-to-substrate adhesion, thus uncovering an unconventional modifier of HD pathology.
Keywords: CAG-repeat expansion; Huntington's disease; MT: Non-coding RNAs; Neurodegeneration; back-splicing; circRNA.
© 2024 The Author(s).
Conflict of interest statement
J.E.W. serves as a consultant for Laronde. V.C.W. was a founding scientific advisory board member with financial interest in Triplet Therapeutics Inc. Her financial interests were reviewed and are managed by Massachusetts General Hospital and Mass General Brigham in accordance with their conflict-of-interest policies. She is a scientific advisory board member of LoQus23 Therapeutics Ltd. and has provided paid consulting services to Acadia Pharmaceuticals Inc., Alnylam Inc., Biogen Inc., Passage Bio, and Rgenta Therapeutics. She has received research support from Pfizer Inc.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
