

Formation of the pyruvoyl-dependent proline reductase Prd from Clostridioides difficile requires the maturation enzyme PrdH
- PMID: 38979079
- PMCID: PMC11229817
- DOI: 10.1093/pnasnexus/pgae249
Formation of the pyruvoyl-dependent proline reductase Prd from Clostridioides difficile requires the maturation enzyme PrdH
Abstract
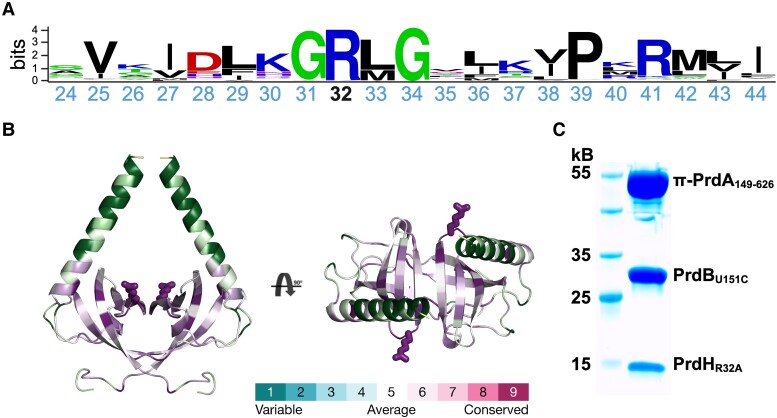
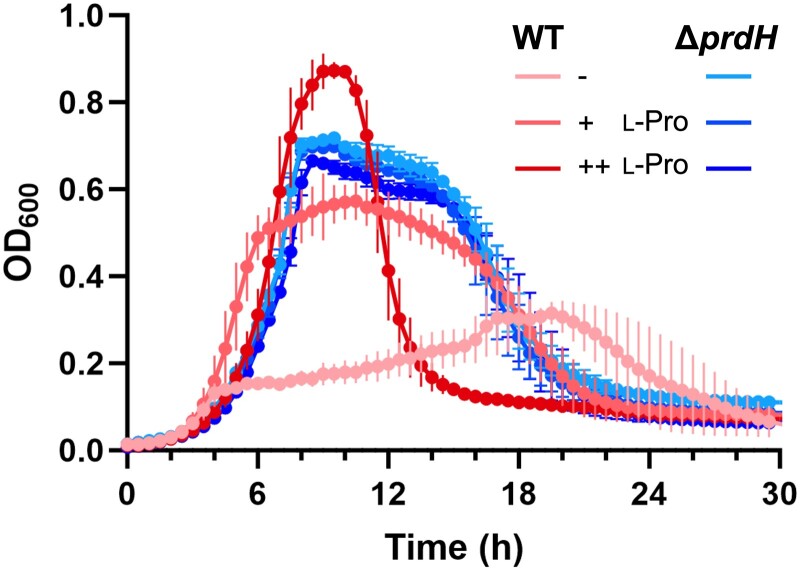
Stickland fermentation, the coupled oxidation and reduction of amino acid pairs, is a major pathway for obtaining energy in the nosocomial bacterium Clostridioides difficile. D-proline is the preferred substrate for the reductive path, making it not only a key component of the general metabolism but also impacting on the expression of the clostridial toxins TcdA and TcdB. D-proline reduction is catalyzed by the proline reductase Prd, which belongs to the pyruvoyl-dependent enzymes. These enzymes are translated as inactive proenzymes and require subsequent processing to install the covalently bound pyruvate. Whereas pyruvoyl formation by intramolecular serinolysis has been studied in unrelated enzymes, details about pyruvoyl generation by cysteinolysis as in Prd are lacking. Here, we show that Prd maturation requires a small dimeric protein that we have named PrdH. PrdH (CD630_32430) is co-encoded with the PrdA and PrdB subunits of Prd and also found in species producing similar reductases. By producing stable variants of PrdA and PrdB, we demonstrate that PrdH-mediated cleavage and pyruvoyl formation in the PrdA subunit requires PrdB, which can be harnessed to produce active recombinant Prd for subsequent analyses. We further created PrdA- and PrdH-mutants to get insight into the interaction of the components and into the processing reaction itself. Finally, we show that deletion of prdH renders C. difficile insensitive to proline concentrations in culture media, suggesting that this processing factor is essential for proline utilization. Due to the link between Stickland fermentation and pathogenesis, we suggest PrdH may be an attractive target for drug development.
Keywords: Clostridioides difficile; Stickland metabolism; autoproteolysis; proline reductase; pyruvoyl enzyme.
© The Author(s) 2024. Published by Oxford University Press on behalf of National Academy of Sciences.
Figures






References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
