

This is a preprint.
A broadly-neutralizing antibody against Ebolavirus glycoprotein that potentiates the breadth and neutralization potency of other antibodies
- PMID: 38979279
- PMCID: PMC11230233
- DOI: 10.1101/2024.06.21.600001
A broadly-neutralizing antibody against Ebolavirus glycoprotein that potentiates the breadth and neutralization potency of other antibodies
Abstract
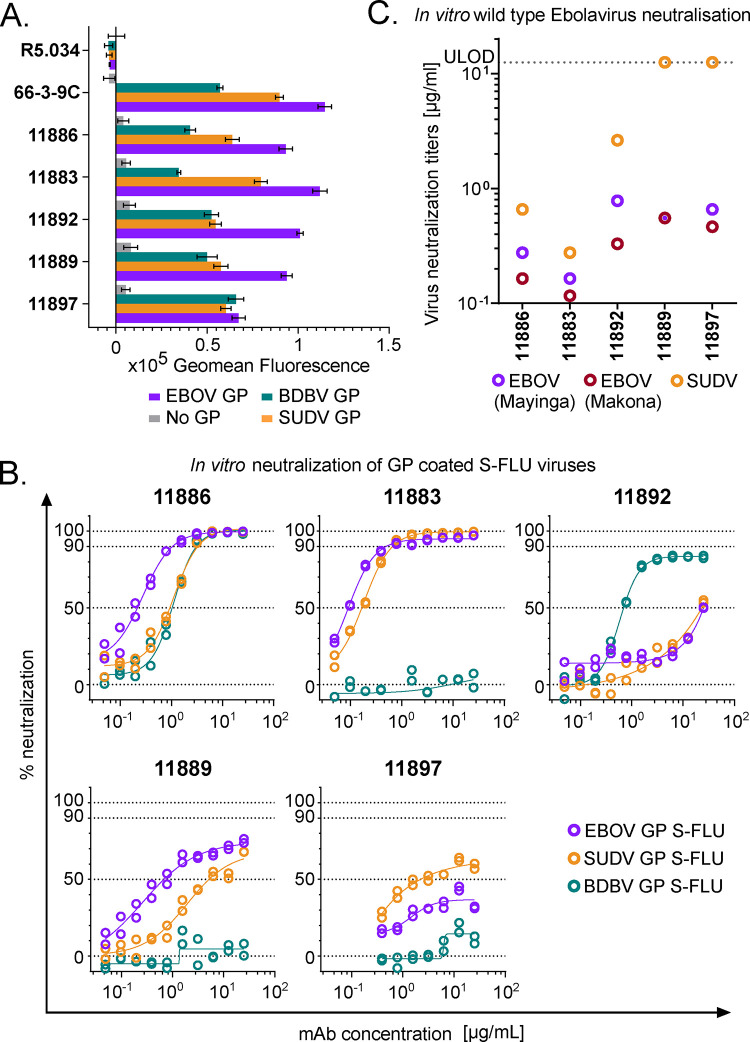
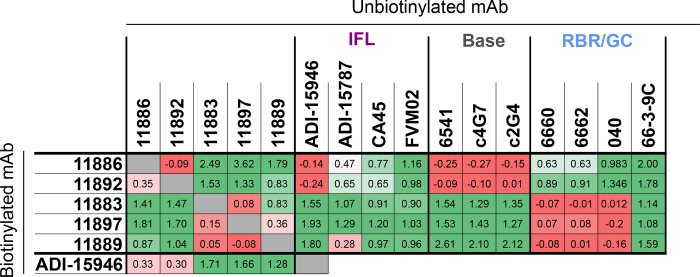
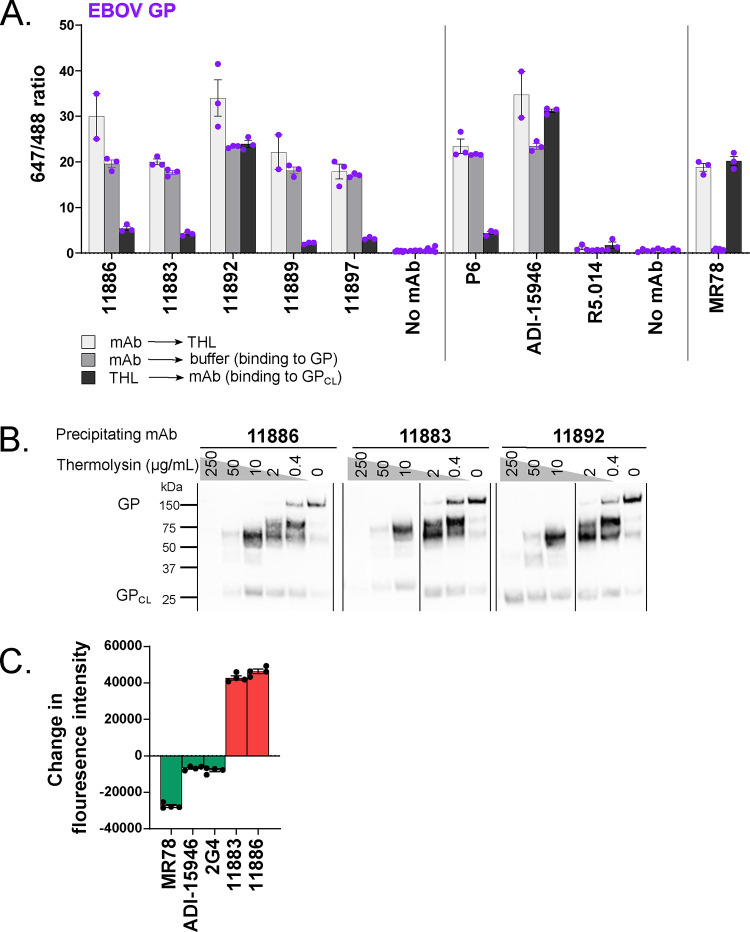
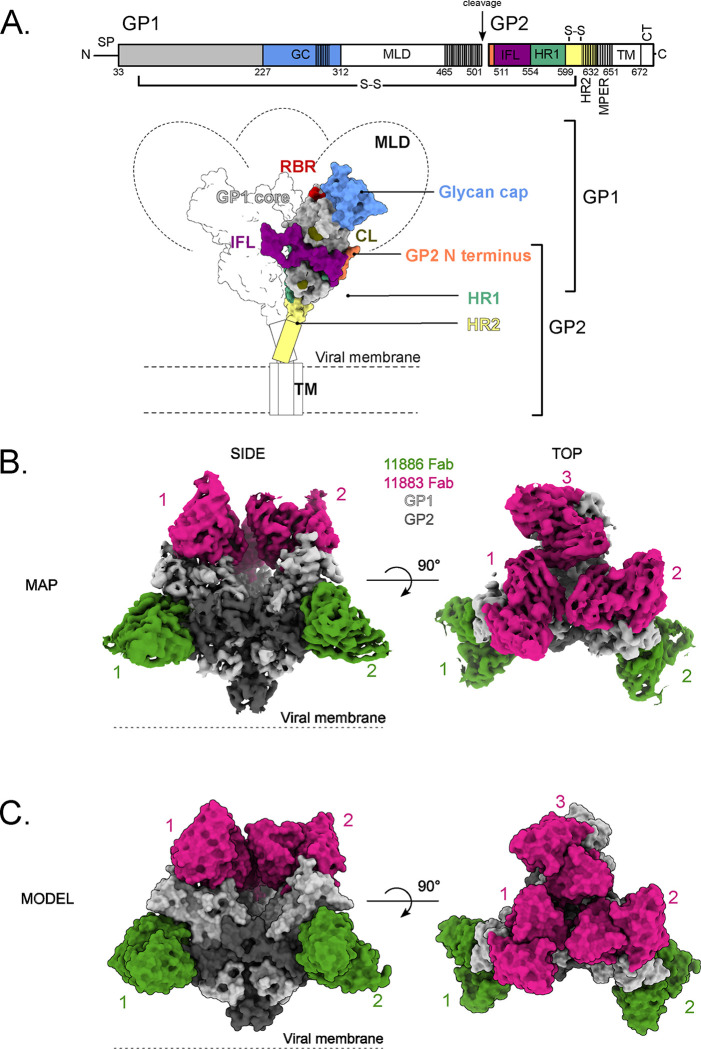
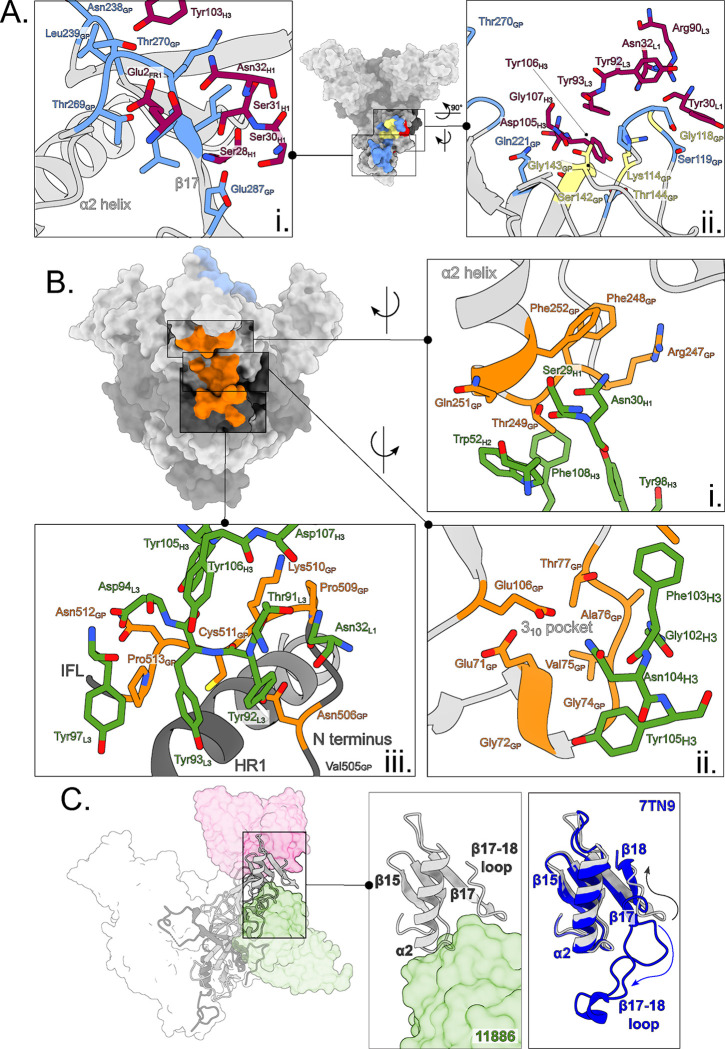
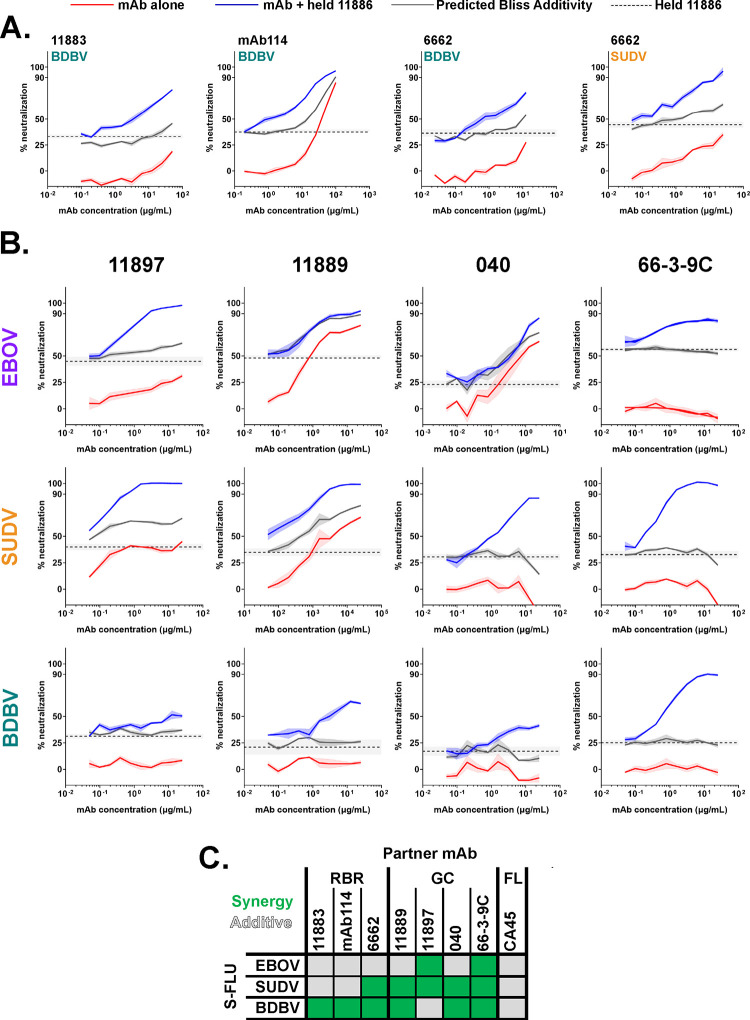
Ebolavirus disease (EVD) is caused by multiple species of Ebolavirus. Monoclonal antibodies (mAbs) against the virus glycoprotein (GP) are the only class of therapeutic approved for treatment of EVD caused by Zaire ebolavirus (EBOV). Therefore, mAbs targeting multiple Ebolavirus species may represent the next generation of EVD therapeutics. Broadly reactive anti-GP mAbs were produced; among these, mAbs 11886 and 11883 were broadly neutralizing in vitro. A 3.0 Å cryo-electron microscopy structure of EBOV GP bound to both mAbs shows that 11886 binds a novel epitope bridging the glycan cap (GC), 310 pocket and GP2 N-terminus, whereas 11883 binds the receptor binding region (RBR) and GC. In vitro, 11886 synergized with a range of mAbs with epitope specificities spanning the RBR/GC, including 11883. Notably, 11886 increased the breadth of neutralization by partner mAbs against different Ebolavirus species. These data provide a strategic route to design improved mAb-based next-generation EVD therapeutics.
Conflict of interest statement
Declaration of Interests DJL, VO’D are employees of UCB Pharma. DJL holds stock and/or stock options in UCB Pharma. FRD, SJD, DJL, VO’D, ART, PR and LS are inventors on patent applications relating to anti-Ebolavirus GP antibodies.
Figures
References
-
- Breman J.G., Heymann D.L., Lloyd G., Mccormick J.B., Miatudila M., Murphy F.A., Muyembé-Tamfun J.-J., Piot P., Ruppol J.-F., Sureau P., et al. (2016). Discovery and Description of Ebola Zaire Virus in 1976 and Relevance to the West African Epidemic During 2013–2016. Journal of Infectious Diseases 214, S93–S101. 10.1093/infdis/jiw207. - DOI - PMC - PubMed
-
- Formenty P., Hatz C., Le Guenno B., Stoll A., Rogenmoser P., and Widmer A. (1999). Human Infection Due to Ebola Virus, Subtype Côte d’Ivoire Clinical and Biologic Presentation. The Journal of Infectious Diseases. - PubMed
-
- Towner J.S., Sealy T.K., Khristova M.L., Albariño C.G., Conlan S., Reeder S.A., Quan P.L., Lipkin W.I., Downing R., Tappero J.W., et al. (2008). Newly discovered ebola virus associated with hemorrhagic fever outbreak in Uganda. PLoS Pathog 4, e1000212. 10.1371/journal.ppat.1000212. - DOI - PMC - PubMed
-
- Goldstein T., Anthony S.J., Gbakima A., Bird B.H., Bangura J., Tremeau-Bravard A., Belaganahalli M.N., Wells H.L., Dhanota J.K., Liang E., et al. (2018). The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat Microbiol 3, 1084–1089. 10.1038/s41564-018-0227-2. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous