

IL-27 disturbs lipid metabolism and restrains mitochondrial activity to inhibit γδ T17 cell-mediated skin inflammation
- PMID: 38982043
- PMCID: PMC11233514
- DOI: 10.1038/s41419-024-06887-0
IL-27 disturbs lipid metabolism and restrains mitochondrial activity to inhibit γδ T17 cell-mediated skin inflammation
Abstract
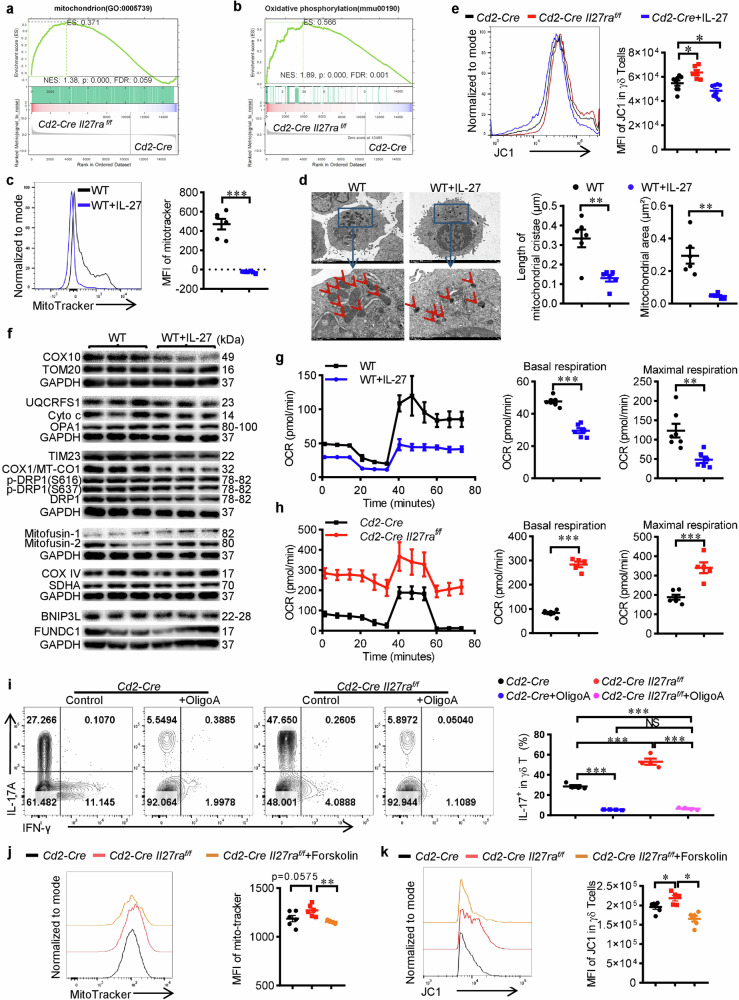
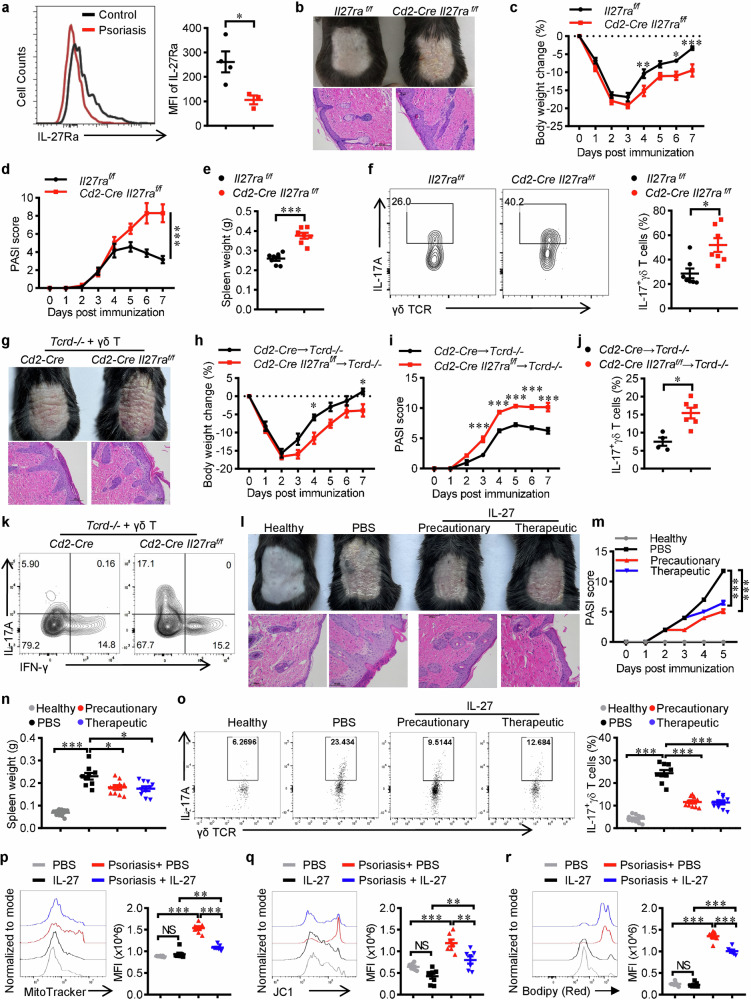
IL-17+ γδ T cells (γδ T17) are kick-starters of inflammation due to their strict immunosurveillance of xenobiotics or cellular damages and rapid response to pro-inflammatory stimulators. IL-27 is a well-recognized pleiotropic immune regulator with potent inhibitory effects on type 17 immune responses. However, its actions on γδ T17 mediated inflammation and the underlying mechanisms are less well understood. Here we find that IL-27 inhibits the production of IL-17 from γδ T cells. Mechanistically, IL-27 promotes lipolysis while inhibits lipogenesis, thus reduces the accumulation of lipids and subsequent membrane phospholipids, which leads to mitochondrial deactivation and ensuing reduction of IL-17. More importantly, Il27ra deficient γδ T cells are more pathogenic in an imiquimod-induced murine psoriasis model, while intracutaneous injection of rmIL-27 ameliorates psoriatic inflammation. In summary, this work uncovered the metabolic basis for the immune regulatory activity of IL-27 in restraining γδ T17 mediated inflammation, which provides novel insights into IL-27/IL-27Ra signaling, γδ T17 biology and the pathogenesis of psoriasis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
- 2023B0303000004/Guangdong Science and Technology Department (Science and Technology Department, Guangdong Province)
- 2022A1515111130/Guangdong Science and Technology Department (Science and Technology Department, Guangdong Province)
- 2023B0303000004/Guangdong Science and Technology Department (Science and Technology Department, Guangdong Province)
- 2023B1515020011/Guangdong Science and Technology Department (Science and Technology Department, Guangdong Province)
- 82300952/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
