

Composite branched and linear F-actin maximize myosin-induced membrane shape changes in a biomimetic cell model
- PMID: 38987288
- PMCID: PMC11236970
- DOI: 10.1038/s42003-024-06528-4
Composite branched and linear F-actin maximize myosin-induced membrane shape changes in a biomimetic cell model
Abstract
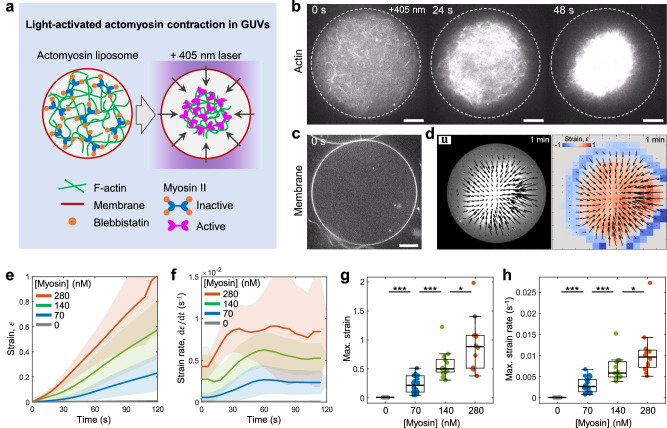
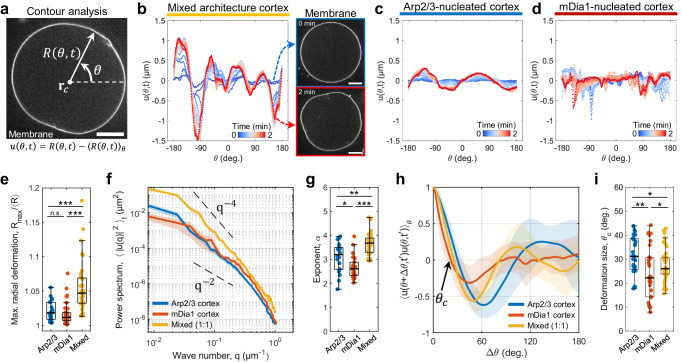
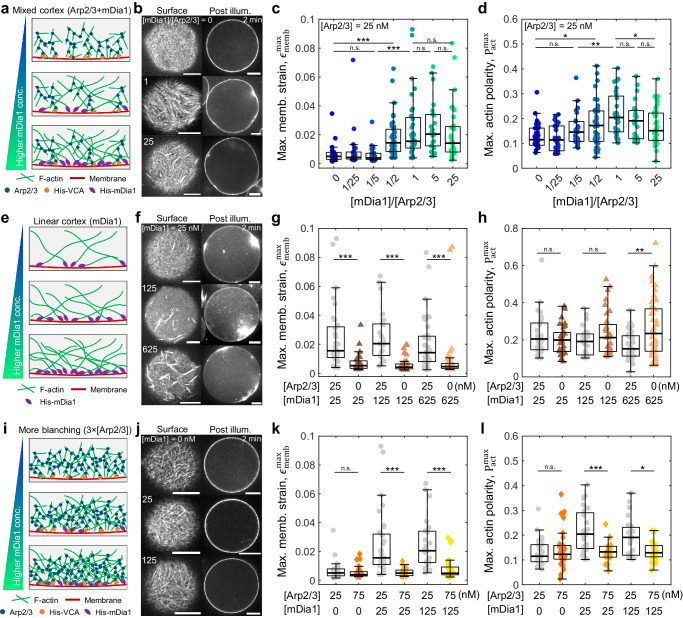
The architecture of the actin cortex determines the generation and transmission of stresses, during key events from cell division to migration. However, its impact on myosin-induced cell shape changes remains unclear. Here, we reconstitute a minimal model of the actomyosin cortex with branched or linear F-actin architecture within giant unilamellar vesicles (GUVs, liposomes). Upon light activation of myosin, neither the branched nor linear F-actin architecture alone induces significant liposome shape changes. The branched F-actin network forms an integrated, membrane-bound "no-slip boundary" -like cortex that attenuates actomyosin contractility. By contrast, the linear F-actin network forms an unintegrated "slip boundary" -like cortex, where actin asters form without inducing membrane deformations. Notably, liposomes undergo significant deformations at an optimized balance of branched and linear F-actin networks. Our findings highlight the pivotal roles of branched F-actin in force transmission and linear F-actin in force generation to yield membrane shape changes.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
