

Systematic optimization of prime editing for the efficient functional correction of CFTR F508del in human airway epithelial cells
- PMID: 38987629
- PMCID: PMC11754097
- DOI: 10.1038/s41551-024-01233-3
Systematic optimization of prime editing for the efficient functional correction of CFTR F508del in human airway epithelial cells
Abstract
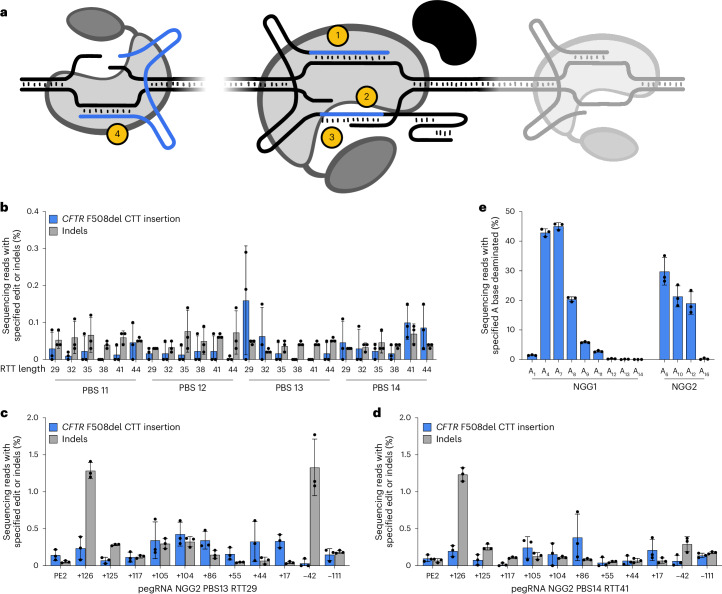
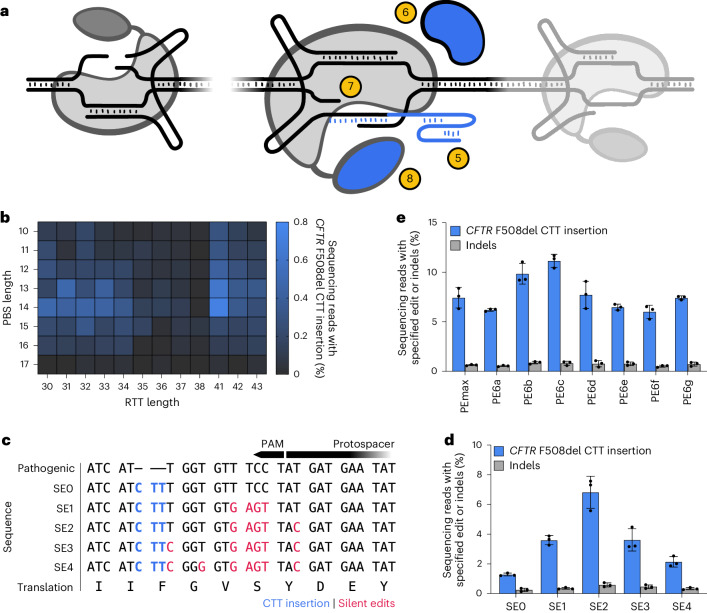
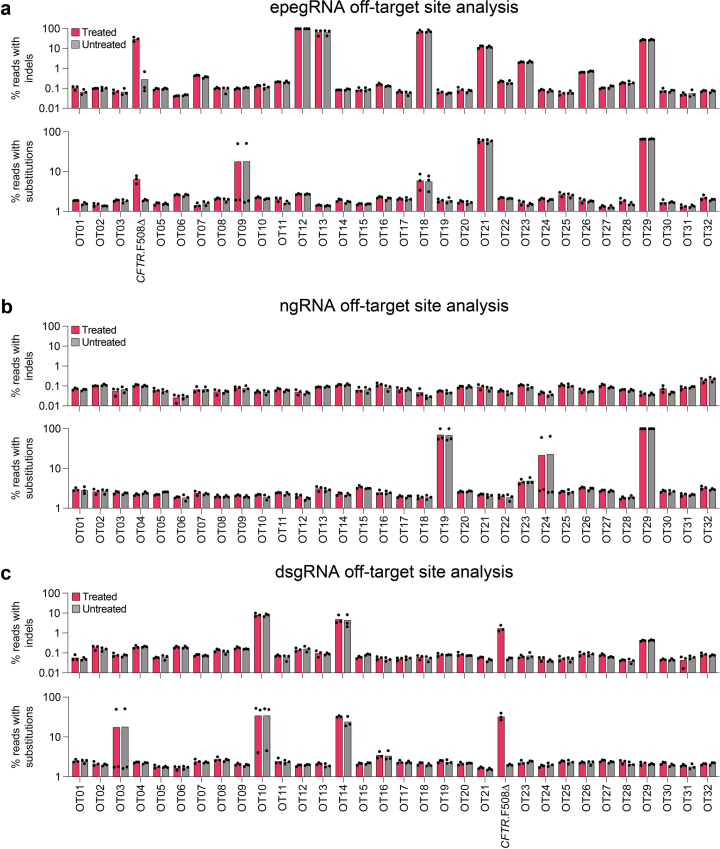
Prime editing (PE) enables precise and versatile genome editing without requiring double-stranded DNA breaks. Here we describe the systematic optimization of PE systems to efficiently correct human cystic fibrosis (CF) transmembrane conductance regulator (CFTR) F508del, a three-nucleotide deletion that is the predominant cause of CF. By combining six efficiency optimizations for PE-engineered PE guide RNAs, the PEmax architecture, the transient expression of a dominant-negative mismatch repair protein, strategic silent edits, PE6 variants and proximal 'dead' single-guide RNAs-we increased correction efficiencies for CFTR F508del from less than 0.5% in HEK293T cells to 58% in immortalized bronchial epithelial cells (a 140-fold improvement) and to 25% in patient-derived airway epithelial cells. The optimizations also resulted in minimal off-target editing, in edit-to-indel ratios 3.5-fold greater than those achieved by nuclease-mediated homology-directed repair, and in the functional restoration of CFTR ion channels to over 50% of wild-type levels (similar to those achieved via combination treatment with elexacaftor, tezacaftor and ivacaftor) in primary airway cells. Our findings support the feasibility of a durable one-time treatment for CF.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: A.A.S., C.H. and D.R.L. have filed patent applications on prime editing through the Broad Institute. P.B.M.J. is on the supervisory advisory board and performs sponsored research for Spirovant Sciences, Inc. D.R.L. is a consultant and equity owner of Prime Medicine, Beam Therapeutics, Pairwise Plants, Exo Therapeutics, Nvelop Therapeutics and Chroma Medicine, all companies that use or deliver genome editing or epigenome-modulating agents. The other authors declare no competing interests.
Figures











References
MeSH terms
Substances
Grants and funding
- K99 HL163805/HL/NHLBI NIH HHS/United States
- P01 HL152960/HL/NHLBI NIH HHS/United States
- RM1 HG009490/HG/NHGRI NIH HHS/United States
- R00 HL163805/HL/NHLBI NIH HHS/United States
- P01HL152960/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R35GM118062/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R35 GM118062/GM/NIGMS NIH HHS/United States
- Liu investigatorship/Howard Hughes Medical Institute (HHMI)
- T32 GM008313/GM/NIGMS NIH HHS/United States
- UH3 HL147366/HL/NHLBI NIH HHS/United States
- RM1HG009490/U.S. Department of Health & Human Services | NIH | National Human Genome Research Institute (NHGRI)
- U24 HG010423/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
