

Macrophage-derived exosomes promote telomere fragility and senescence in tubular epithelial cells by delivering miR-155
- PMID: 38987851
- PMCID: PMC11238407
- DOI: 10.1186/s12964-024-01708-5
Macrophage-derived exosomes promote telomere fragility and senescence in tubular epithelial cells by delivering miR-155
Abstract
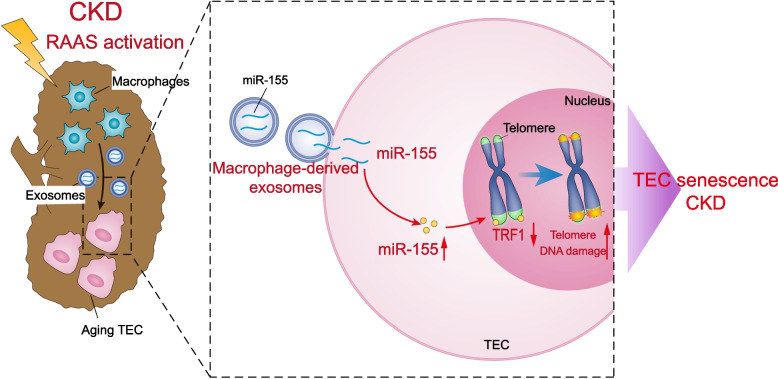
Background: Chronic kidney disease (CKD) is highly prevalent worldwide, and its global burden is substantial and growing. CKD displays a number of features of accelerated senescence. Tubular cell senescence is a common biological process that contributes to CKD progression. Tubulointerstitial inflammation is a driver of tubular cell senescence and a common characteristic of CKD. However, the mechanism by which the interstitial inflammation drives tubular cell senescence remains unclear. This paper aims to explore the role of exosomal miRNAs derived from macrophages in the development of tubular cell senescence.
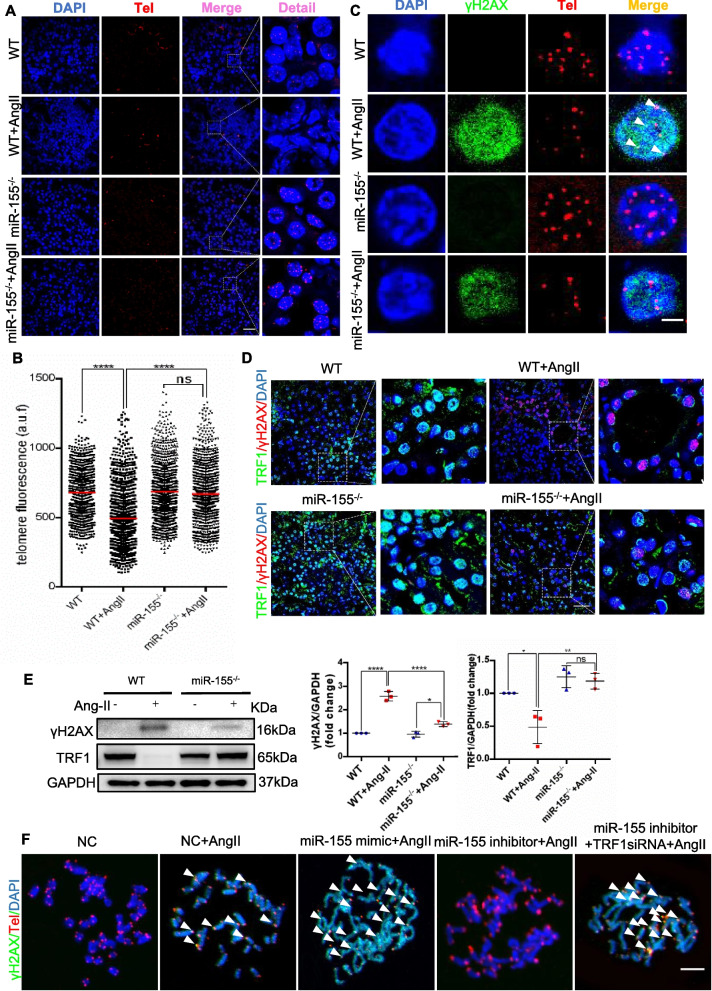
Methods: Among the identified inflammation-related miRNAs, miR-155 is considered to be one of the most important miRNAs involved in the inflammatory response. Macrophages, the primary immune cells that mediate inflammatory processes, contain a high abundance of miR-155 in their released exosomes. We assessed the potential role of miR-155 in tubular cell senescence and renal fibrosis. We subjected miR-155-/- mice and wild-type controls, as well as tubular epithelial cells (TECs), to angiotensin II (AngII)-induced kidney injury. We assessed kidney function and injury using standard techniques. TECs were evaluated for cell senescence and telomere dysfunction in vivo and in vitro. Telomeres were measured by the fluorescence in situ hybridization.
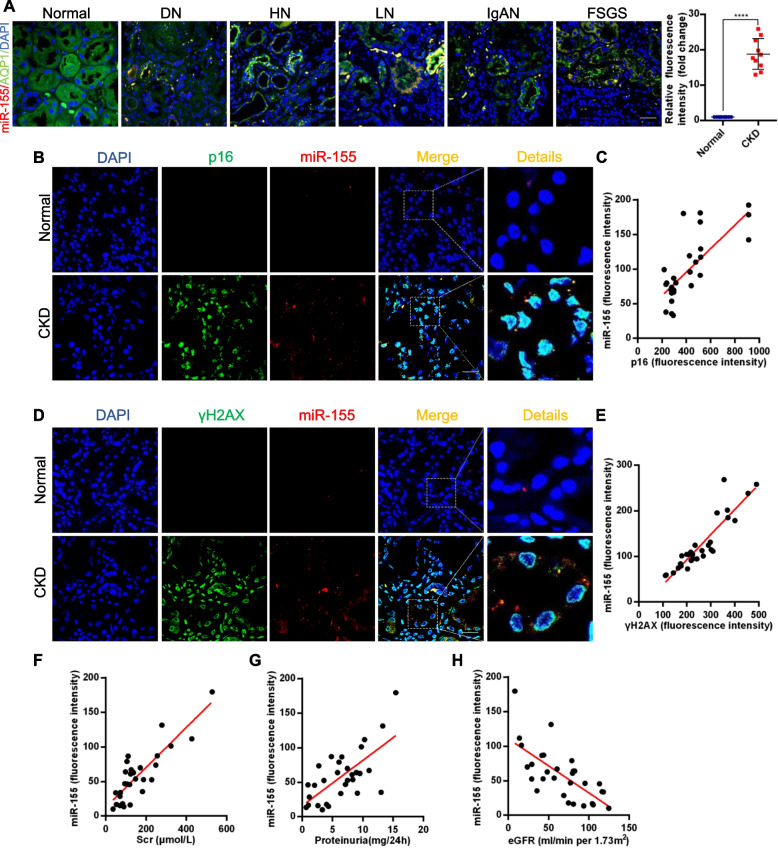
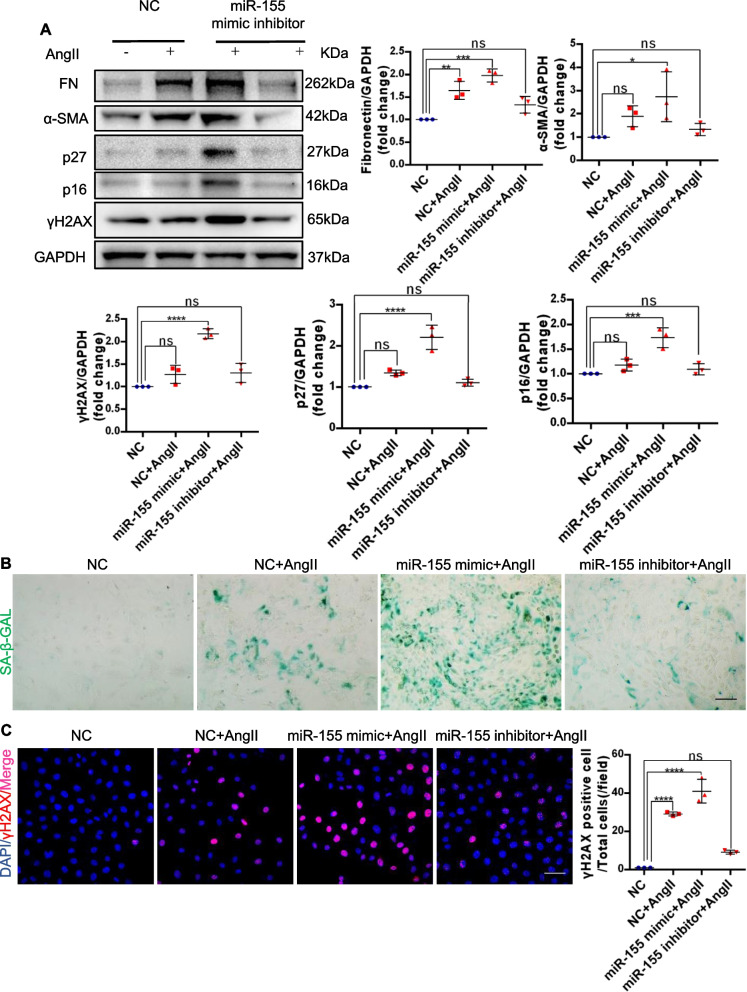
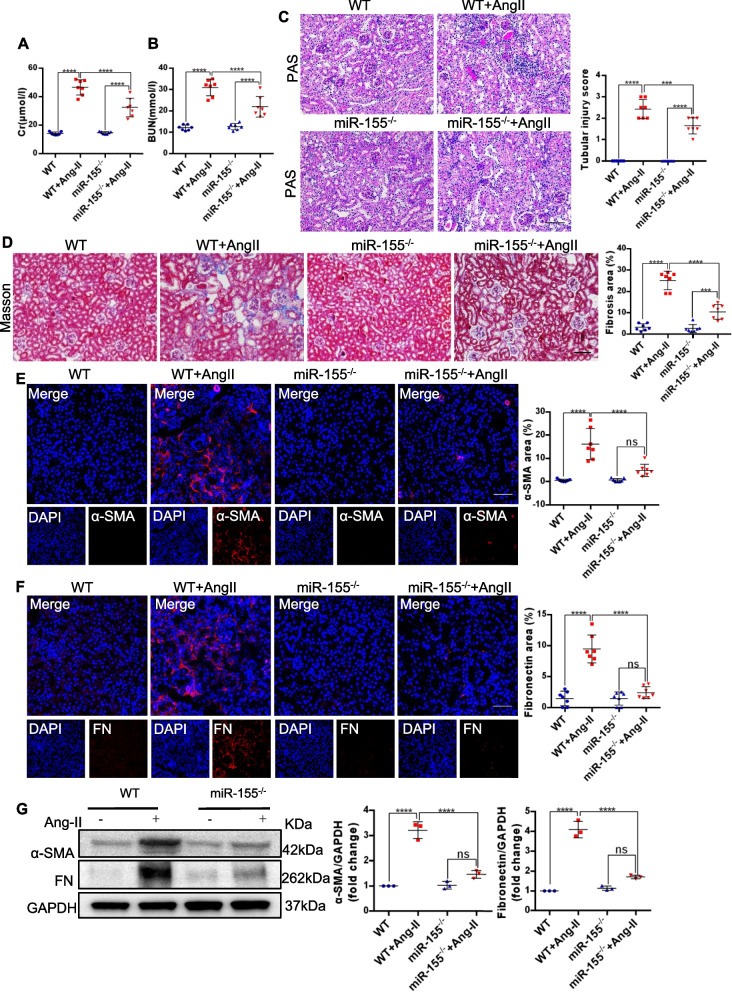
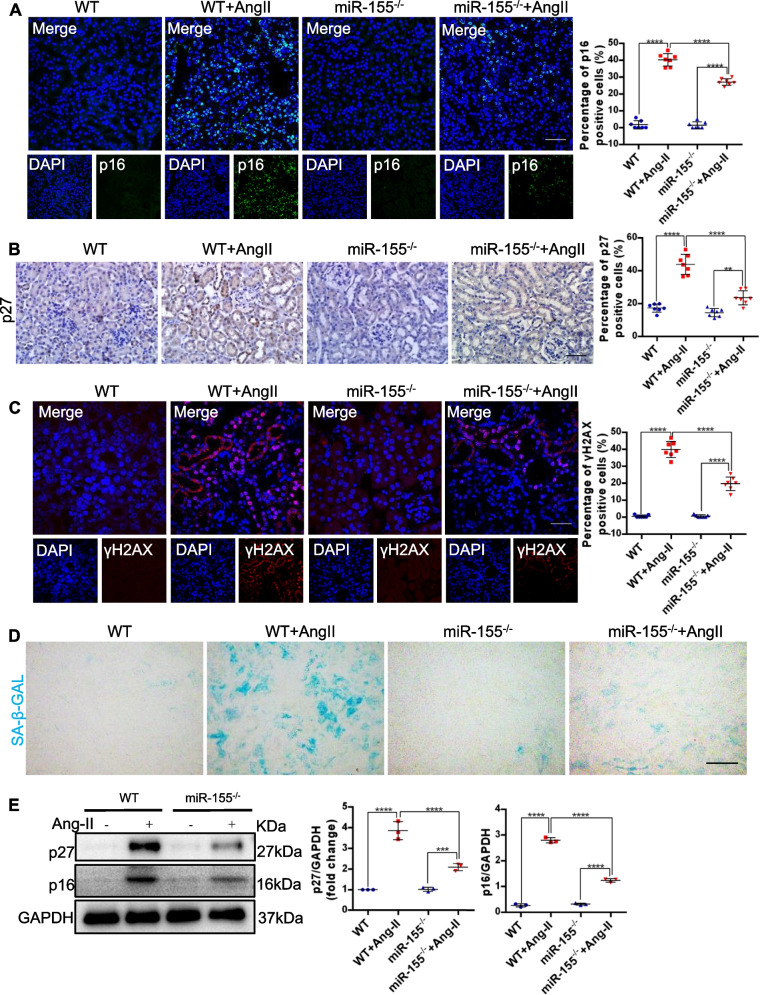
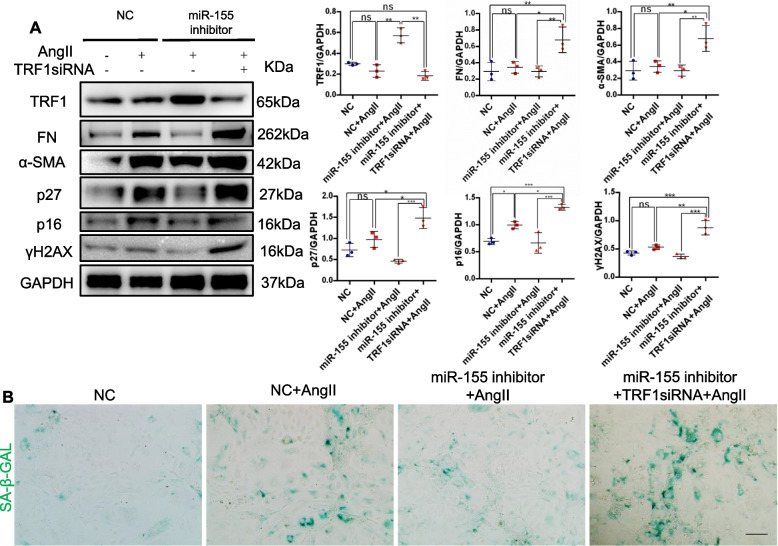
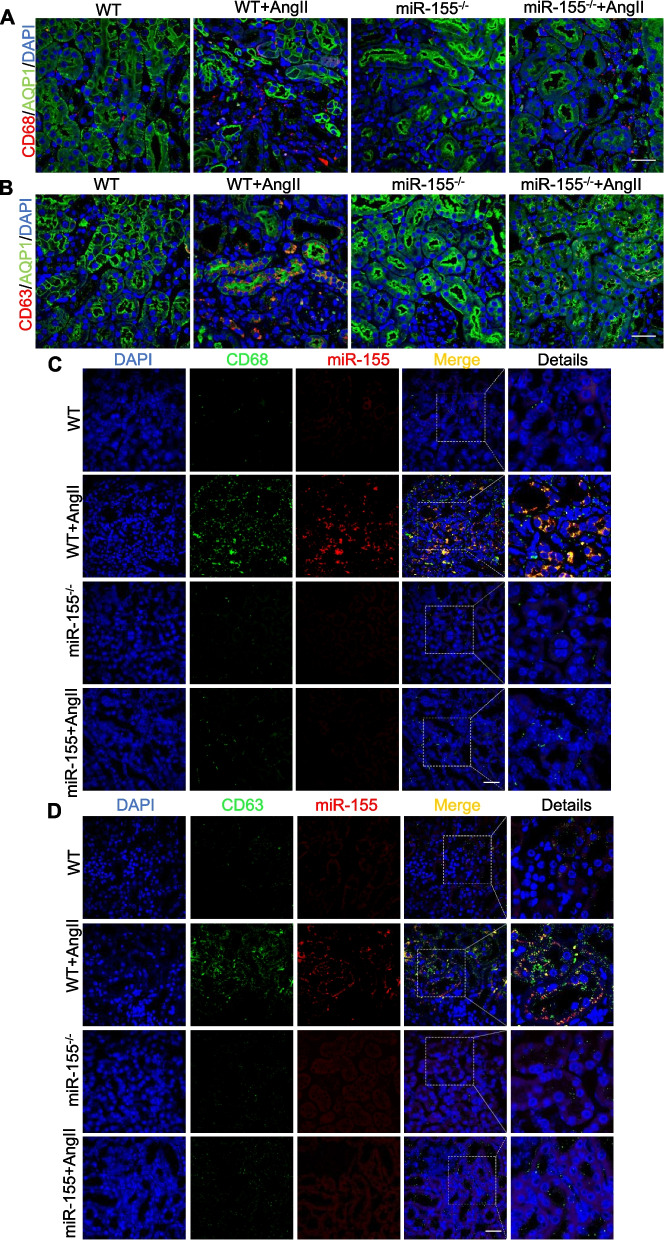
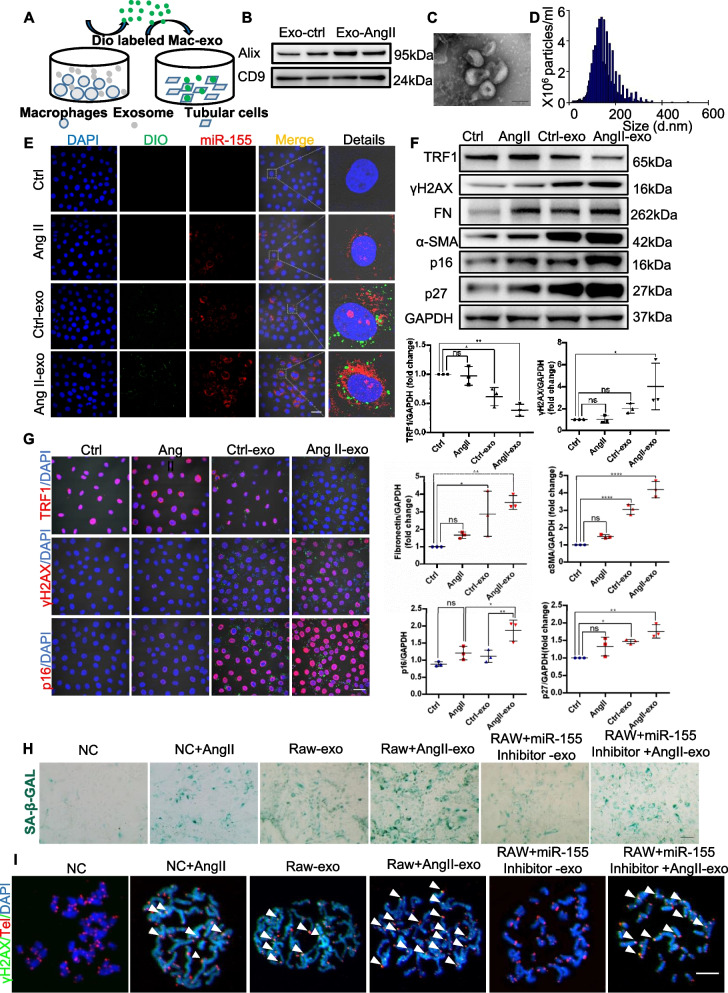
Results: Compared with normal controls, miR-155 was up-regulated in proximal renal tubule cells in CKD patients and mouse models of CKD. Moreover, the expression of miR-155 was positively correlated with the extent of renal fibrosis, eGFR decline and p16INK4A expression. The overexpression of miR-155 exacerbated tubular senescence, evidenced by increased detection of p16INK4A/p21expression and senescence-associated β-galactosidase activity. Notably, miR-155 knockout attenuates renal fibrosis and tubule cell senescence in vivo. Interestingly, once released, macrophages-derived exosomal miR-155 was internalized by TECs, leading to telomere shortening and dysfunction through targeting TRF1. A dual-luciferase reporter assay confirmed that TRF1 was the direct target of miR-155. Thus, our study clearly demonstrates that exosomal miR-155 may mediate communication between macrophages and TECs, subsequently inducing telomere dysfunction and senescence in TECs.
Conclusions: Our work suggests a new mechanism by which macrophage exosomes are involved in the development of tubule senescence and renal fibrosis, in part by delivering miR-155 to target TRF1 to promote telomere dysfunction. Our study may provide novel strategies for the treatment of AngII-induced kidney injury.
Keywords: CKD; Cell senescence; Exosomes; Macrophages; MiR-155; TRF1.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
