

CD74 is a functional MIF receptor on activated CD4+ T cells
- PMID: 38992165
- PMCID: PMC11335222
- DOI: 10.1007/s00018-024-05338-5
CD74 is a functional MIF receptor on activated CD4+ T cells
Abstract
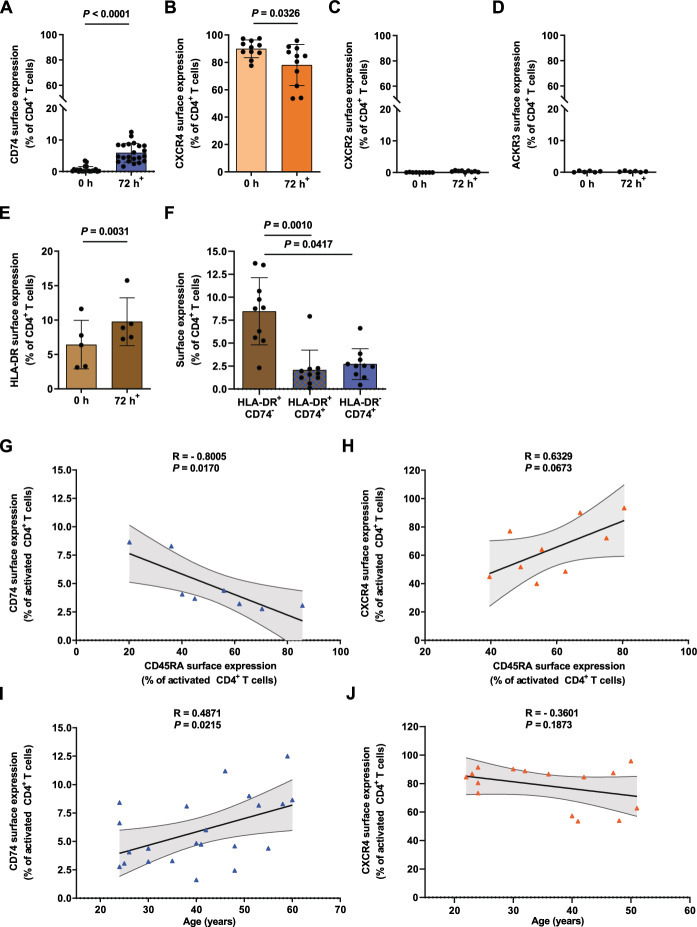
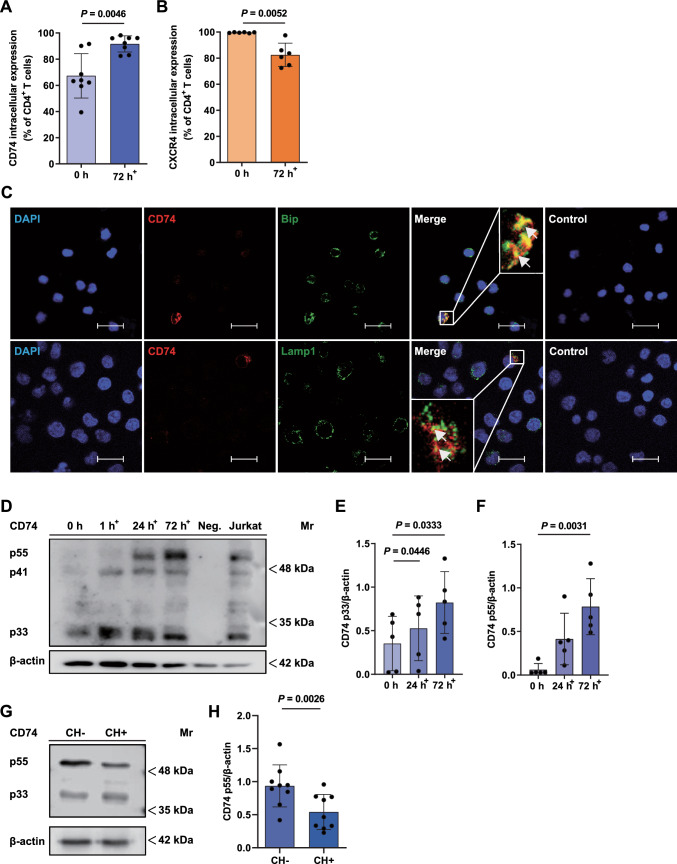
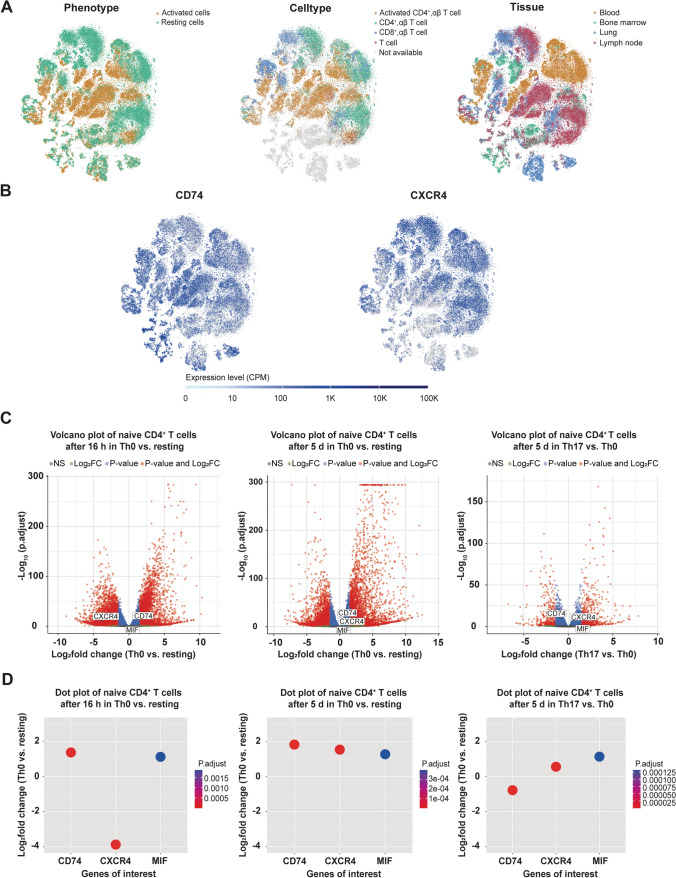
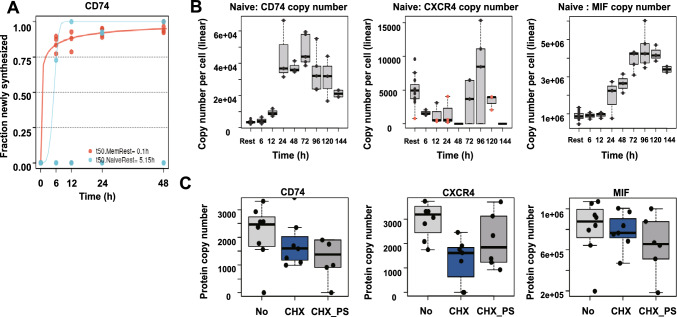
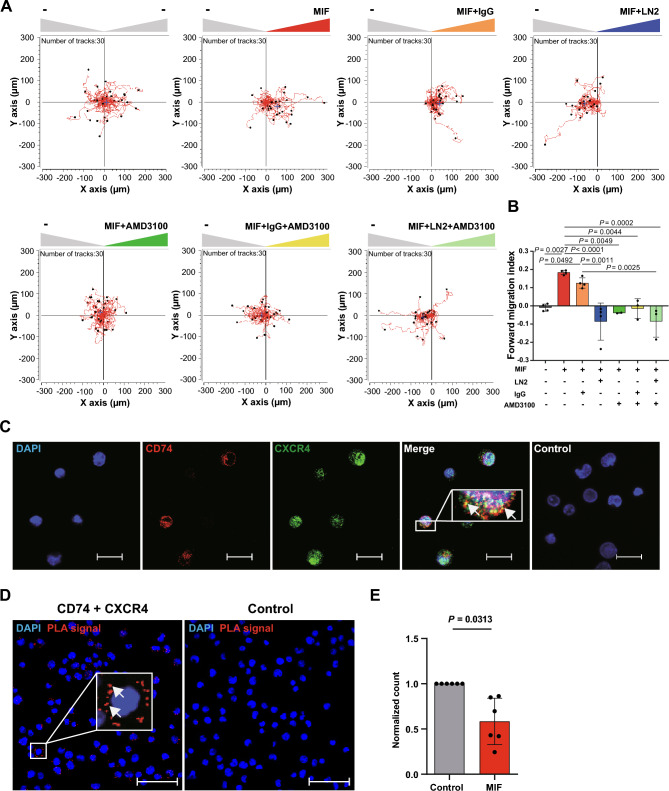
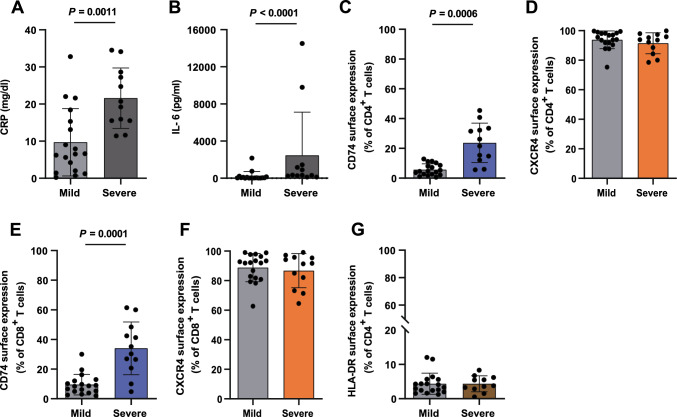
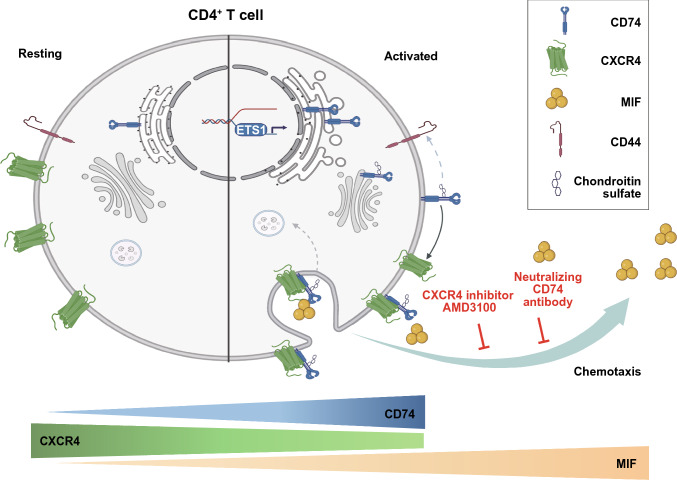
Next to its classical role in MHC II-mediated antigen presentation, CD74 was identified as a high-affinity receptor for macrophage migration inhibitory factor (MIF), a pleiotropic cytokine and major determinant of various acute and chronic inflammatory conditions, cardiovascular diseases and cancer. Recent evidence suggests that CD74 is expressed in T cells, but the functional relevance of this observation is poorly understood. Here, we characterized the regulation of CD74 expression and that of the MIF chemokine receptors during activation of human CD4+ T cells and studied links to MIF-induced T-cell migration, function, and COVID-19 disease stage. MIF receptor profiling of resting primary human CD4+ T cells via flow cytometry revealed high surface expression of CXCR4, while CD74, CXCR2 and ACKR3/CXCR7 were not measurably expressed. However, CD4+ T cells constitutively expressed CD74 intracellularly, which upon T-cell activation was significantly upregulated, post-translationally modified by chondroitin sulfate and could be detected on the cell surface, as determined by flow cytometry, Western blot, immunohistochemistry, and re-analysis of available RNA-sequencing and proteomic data sets. Applying 3D-matrix-based live cell-imaging and receptor pathway-specific inhibitors, we determined a causal involvement of CD74 and CXCR4 in MIF-induced CD4+ T-cell migration. Mechanistically, proximity ligation assay visualized CD74/CXCR4 heterocomplexes on activated CD4+ T cells, which were significantly diminished after MIF treatment, pointing towards a MIF-mediated internalization process. Lastly, in a cohort of 30 COVID-19 patients, CD74 surface expression was found to be significantly upregulated on CD4+ and CD8+ T cells in patients with severe compared to patients with only mild disease course. Together, our study characterizes the MIF receptor network in the course of T-cell activation and reveals CD74 as a novel functional MIF receptor and MHC II-independent activation marker of primary human CD4+ T cells.
Keywords: Atypical chemokine; CD74/invariant chain; CXCR4; MIF; Macrophage migration inhibitory factor; T cells.
© 2024. The Author(s).
Conflict of interest statement
C.S. received speaker honoraria from AstraZeneca on topics outside of the submitted work. J.B. and O.E.B. are inventors on patent applications related to anti-MIF strategies. All other authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
