

Eta-secretase-like processing of the amyloid precursor protein (APP) by the rhomboid protease RHBDL4
- PMID: 38992438
- PMCID: PMC11345391
- DOI: 10.1016/j.jbc.2024.107541
Eta-secretase-like processing of the amyloid precursor protein (APP) by the rhomboid protease RHBDL4
Abstract
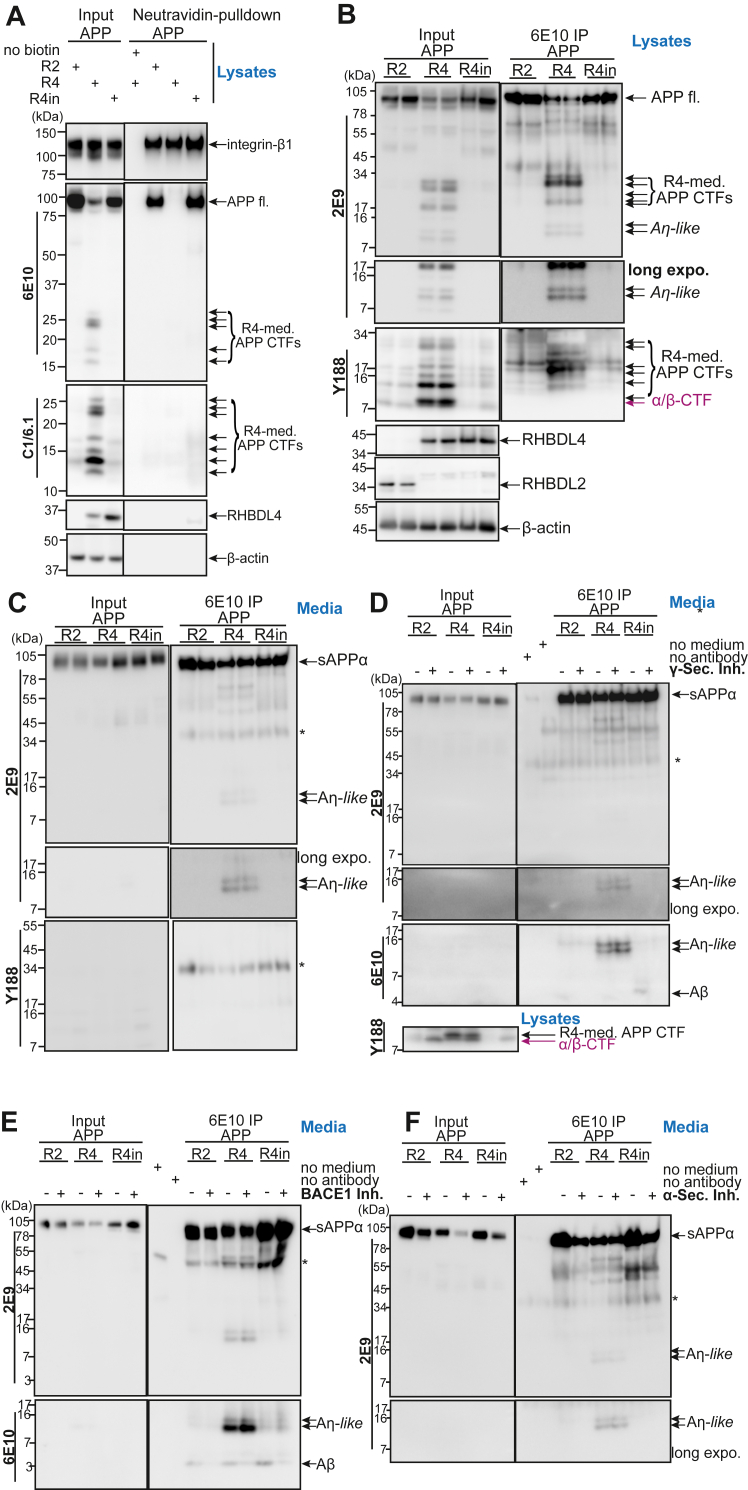
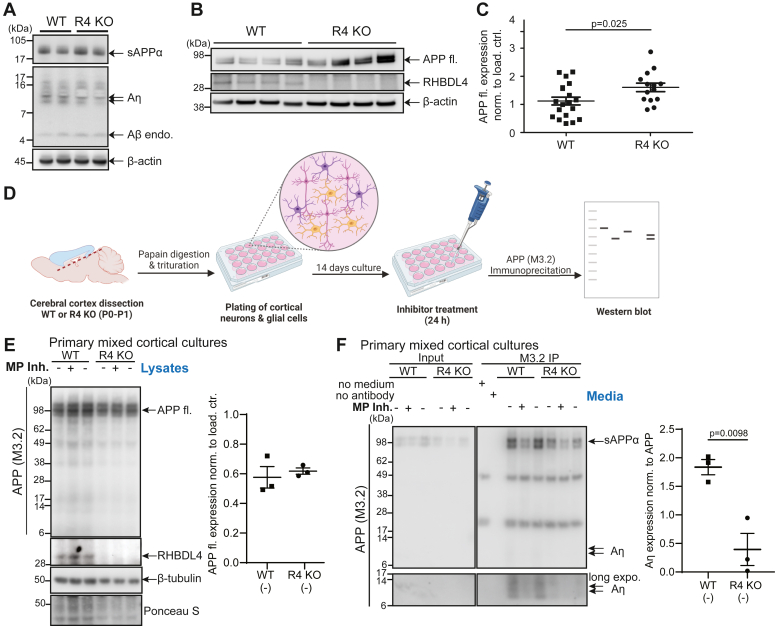
The amyloid precursor protein (APP) is a key protein in Alzheimer's disease synthesized in the endoplasmic reticulum (ER) and translocated to the plasma membrane where it undergoes proteolytic cleavages by several proteases. Conversely, to other known proteases, we previously elucidated rhomboid protease RHBDL4 as a novel APP processing enzyme where several cleavages likely occur already in the ER. Interestingly, the pattern of RHBDL4-derived large APP C-terminal fragments resembles those generated by the η-secretase or MT5-MMP, which was described to generate so-called Aη fragments. The similarity in large APP C-terminal fragments between both proteases raised the question of whether RHBDL4 may contribute to η-secretase activity and Aη-like fragments. Here, we identified two cleavage sites of RHBDL4 in APP by mass spectrometry, which, intriguingly, lie in close proximity to the MT5-MMP cleavage sites. Indeed, we observed that RHBDL4 generates Aη-like fragments in vitro without contributions of α-, β-, or γ-secretases. Such Aη-like fragments are likely generated in the ER since RHBDL4-derived APP-C-terminal fragments do not reach the cell surface. Inherited, familial APP mutations appear to not affect this processing pathway. In RHBDL4 knockout mice, we observed increased cerebral full-length APP in comparison to wild type (WT) in support of RHBDL4 being a physiologically relevant protease for APP. Furthermore, we found secreted Aη fragments in dissociated mixed cortical cultures from WT mice, however significantly fewer Aη fragments in RHBDL4 knockout cultures. Our data underscores that RHBDL4 contributes to the η-secretease-like processing of APP and that RHBDL4 is a physiologically relevant protease for APP.
Keywords: Alzheimer’s disease; Aη; MMP24; MT5-MMP; RHBDD1; RHBDL4; amyloid precursor protein (APP); eta-secretase; rhomboid protease.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
-
- Duran-Aniotz C., Hetz C. Glucose metabolism: a sweet relief of Alzheimer's disease. Curr. Biol. 2016;26:R806–R809. - PubMed
-
- Puglielli L., Tanzi R.E., Kovacs D.M. Alzheimer's disease: the cholesterol connection. Nat. Neurosci. 2003;6:345–351. - PubMed
-
- Spillantini M.G., Goedert M. Tau pathology and neurodegeneration. Lancet Neurol. 2013;12:609–622. - PubMed
-
- Winblad B., Amouyel P., Andrieu S., Ballard C., Brayne C., Brodaty H., et al. Defeating Alzheimer's disease and other dementias: a priority for European science and society. Lancet Neurol. 2016;15:455–532. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
