

A neutral invertase controls cell division besides hydrolysis of sucrose for nutrition during germination and seed setting in rice
- PMID: 38993663
- PMCID: PMC11237924
- DOI: 10.1016/j.isci.2024.110217
A neutral invertase controls cell division besides hydrolysis of sucrose for nutrition during germination and seed setting in rice
Abstract
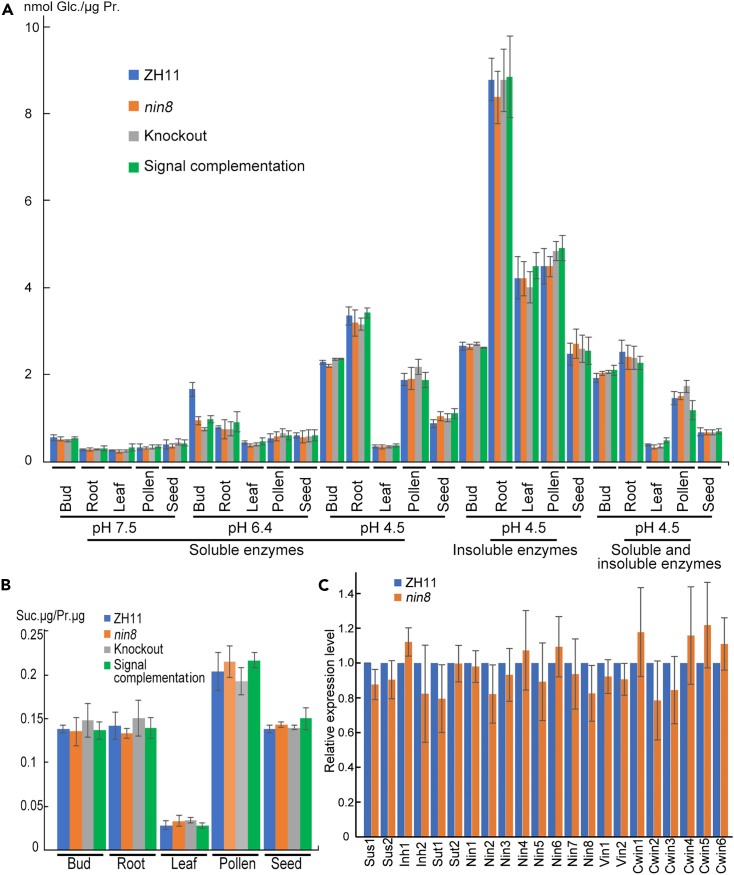
Sucrose is the transport form of carbohydrate in plants serving as signal molecule besides nutrition, but the signaling is elusive. Here, neutral invertase 8 (OsNIN8) mutated at G461R into OsNIN8m, which increased its charge and hydrophobicity, decreased hydrolysis of sucrose to 13% and firmer binding to sucrose than the wildtype. This caused downstream metabolites and energy accumulation forming overnutrition. Paradoxically, division of subinitials in longitudinal cell lineages was only about 15 times but more than 100 times in wildtype, resulting in short radicle. Further, mutation of OsNIN8 into deficiency of hydrolysis but maintenance of sucrose binding allowed cell division until ran out of energy showing the association but not hydrolysis gave the signal. Chemically, sucrose binding to OsNIN8 was exothermic but to OsNIN8m was endothermic. Therefore, OsNIN8m lost the signal function owing to change of thermodynamic state. So, OsNIN8 sensed sucrose for cell division besides hydrolyzed sucrose.
Keywords: Cell biology; Plant biology; Plant development.
© 2024 The Authors.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






References
-
- Humphreys T.E. Sucrose efflux and export from the maize scutellum. Plant Cell Environ. 1987;10:259–266. doi: 10.1111/1365-3040.EP11602277. - DOI
-
- Sánchez-Linares L., Gavilanes-Ruiz M., Diaz-Pontones D., Guzman-Chavez F., Calzada-Alejo V., Zurita-Villegas V., Luna-Loaiza V., Moreno-Sanchez R., Bernal-Lugo I., Sanchez-Nieto S. Early carbon mobilization and radicle protrusion in maize germination. J. Exp. Bot. 2012;63:4513–4526. doi: 10.1093/jxb/ers130. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
