

Evolutionary adaptation of an HP1-protein chromodomain integrates chromatin and DNA sequence signals
- PMID: 38995818
- PMCID: PMC11245307
- DOI: 10.7554/eLife.93194
Evolutionary adaptation of an HP1-protein chromodomain integrates chromatin and DNA sequence signals
Abstract
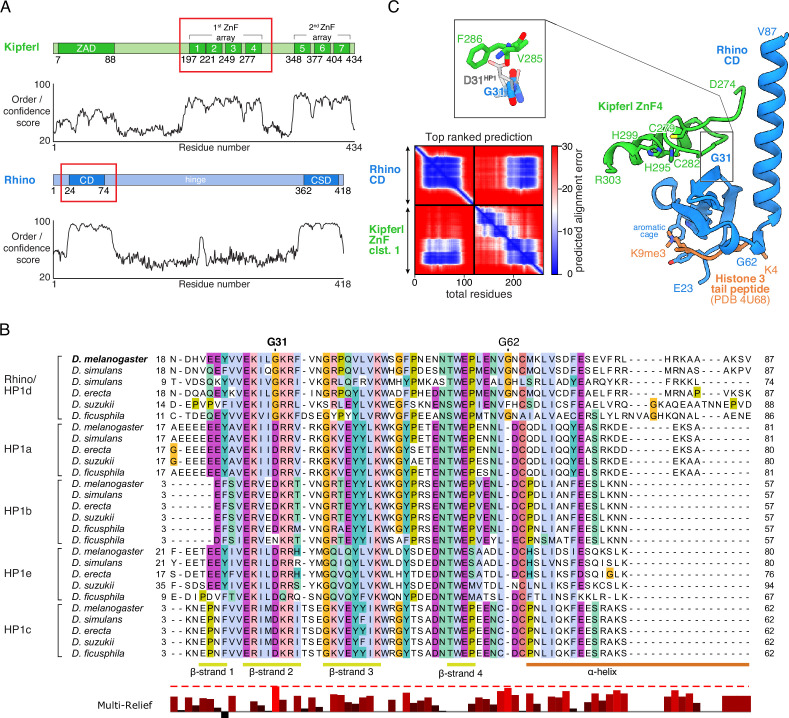
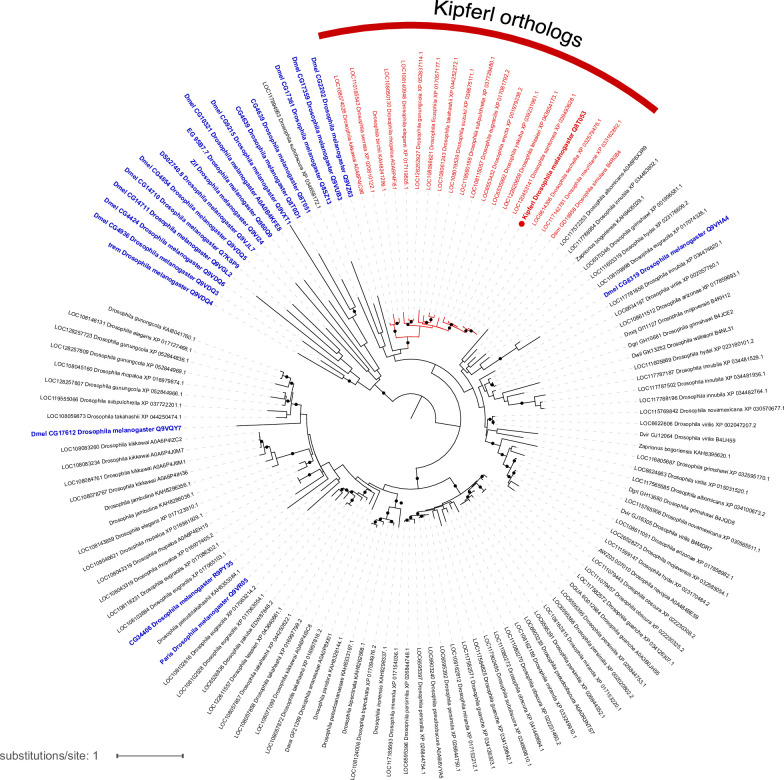
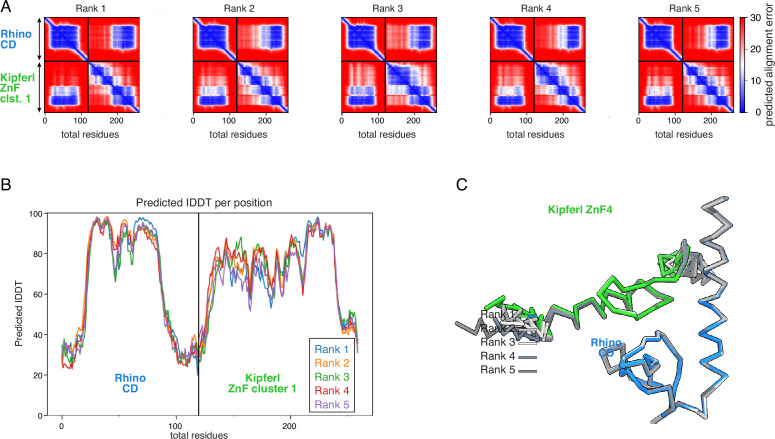
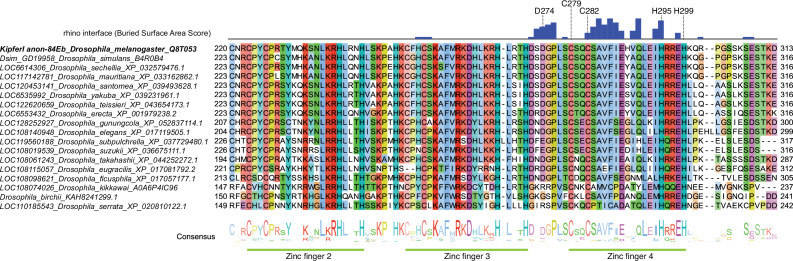
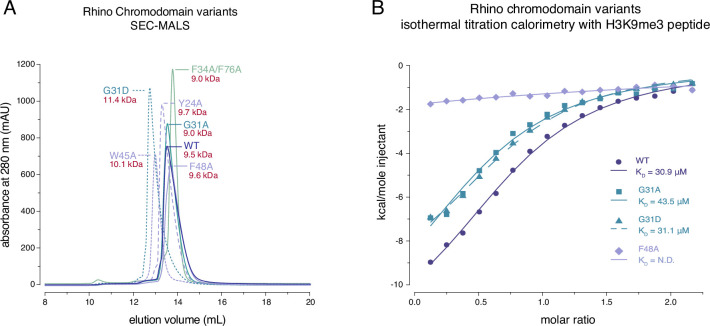
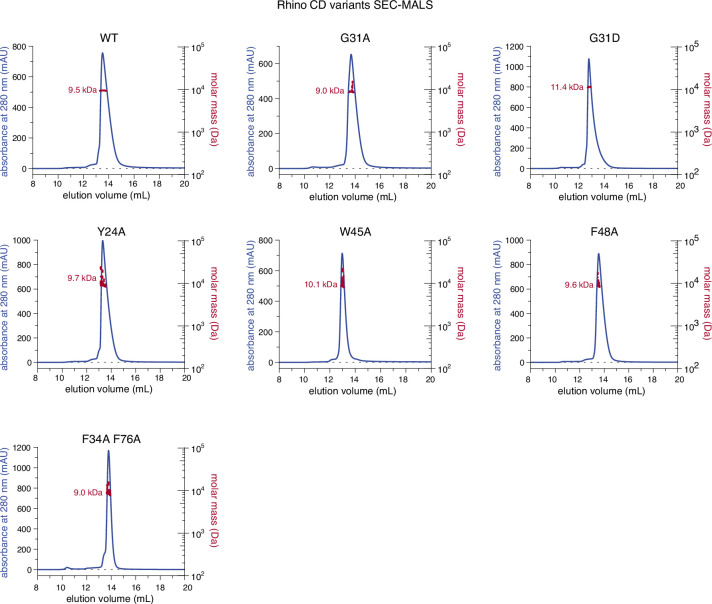
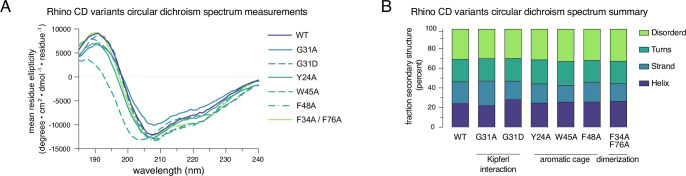
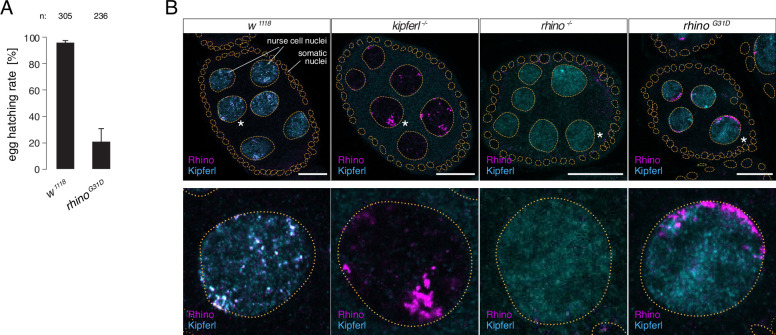
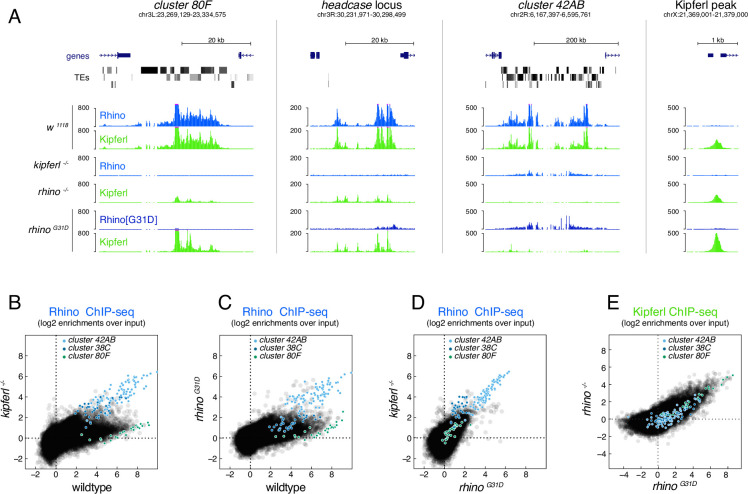
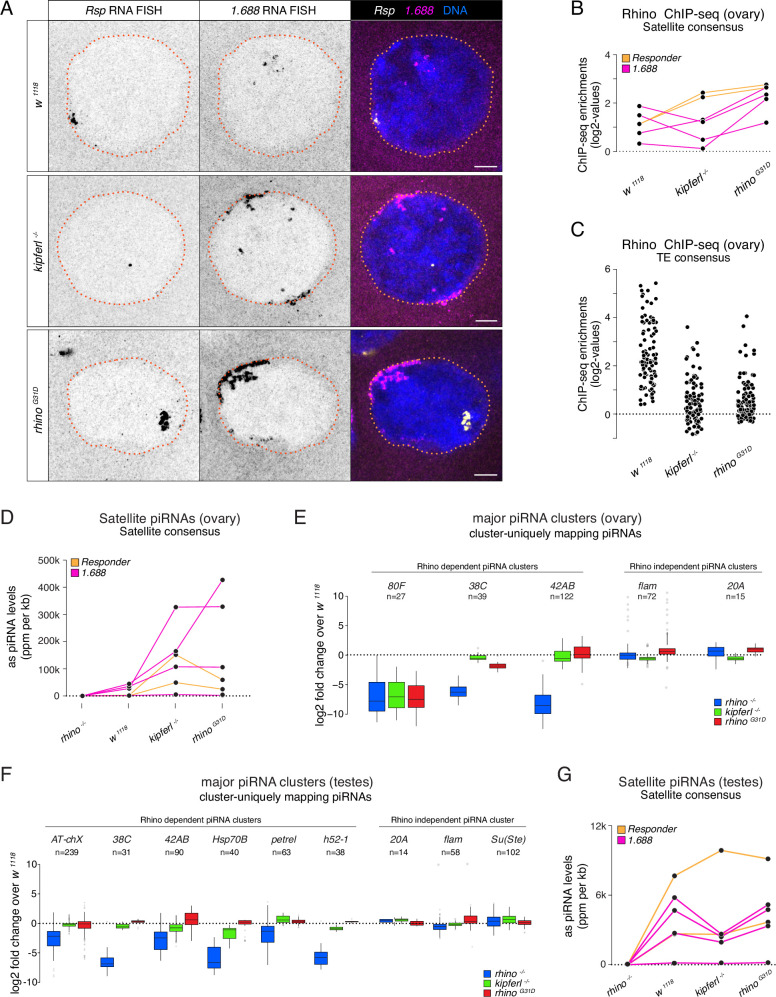
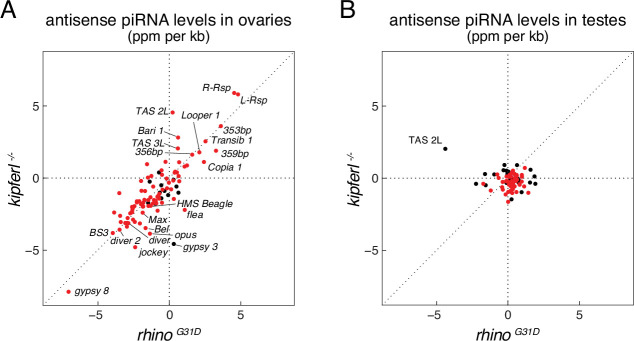
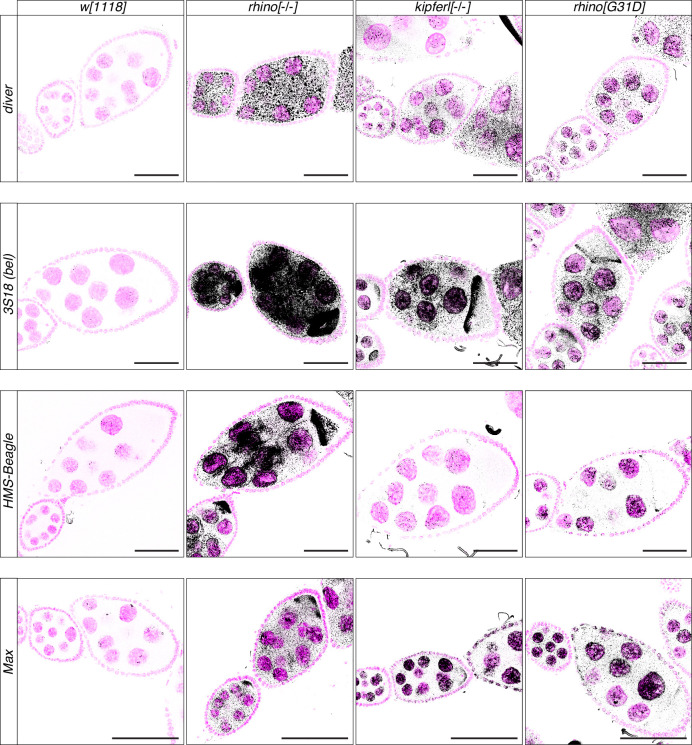
Members of the diverse heterochromatin protein 1 (HP1) family play crucial roles in heterochromatin formation and maintenance. Despite the similar affinities of their chromodomains for di- and tri-methylated histone H3 lysine 9 (H3K9me2/3), different HP1 proteins exhibit distinct chromatin-binding patterns, likely due to interactions with various specificity factors. Previously, we showed that the chromatin-binding pattern of the HP1 protein Rhino, a crucial factor of the Drosophila PIWI-interacting RNA (piRNA) pathway, is largely defined by a DNA sequence-specific C2H2 zinc finger protein named Kipferl (Baumgartner et al., 2022). Here, we elucidate the molecular basis of the interaction between Rhino and its guidance factor Kipferl. Through phylogenetic analyses, structure prediction, and in vivo genetics, we identify a single amino acid change within Rhino's chromodomain, G31D, that does not affect H3K9me2/3 binding but disrupts the interaction between Rhino and Kipferl. Flies carrying the rhinoG31D mutation phenocopy kipferl mutant flies, with Rhino redistributing from piRNA clusters to satellite repeats, causing pronounced changes in the ovarian piRNA profile of rhinoG31D flies. Thus, Rhino's chromodomain functions as a dual-specificity module, facilitating interactions with both a histone mark and a DNA-binding protein.
Keywords: D. melanogaster; HP1 proteins; chromodomain; chromosomes; gene expression; genetics; genomics; heterochromatin; piRNA pathway; transposon silencing; zinc finger protein.
© 2024, Baumgartner et al.
Conflict of interest statement
LB, JI, UH, DH, AS, PD, JB No competing interests declared
Figures

Update of
- doi: 10.1101/2023.09.29.560096
- doi: 10.7554/eLife.93194.1
- doi: 10.7554/eLife.93194.2
References
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
