

Molecular and cellular mechanisms of teneurin signaling in synaptic partner matching
- PMID: 38996528
- PMCID: PMC11833509
- DOI: 10.1016/j.cell.2024.06.022
Molecular and cellular mechanisms of teneurin signaling in synaptic partner matching
Abstract
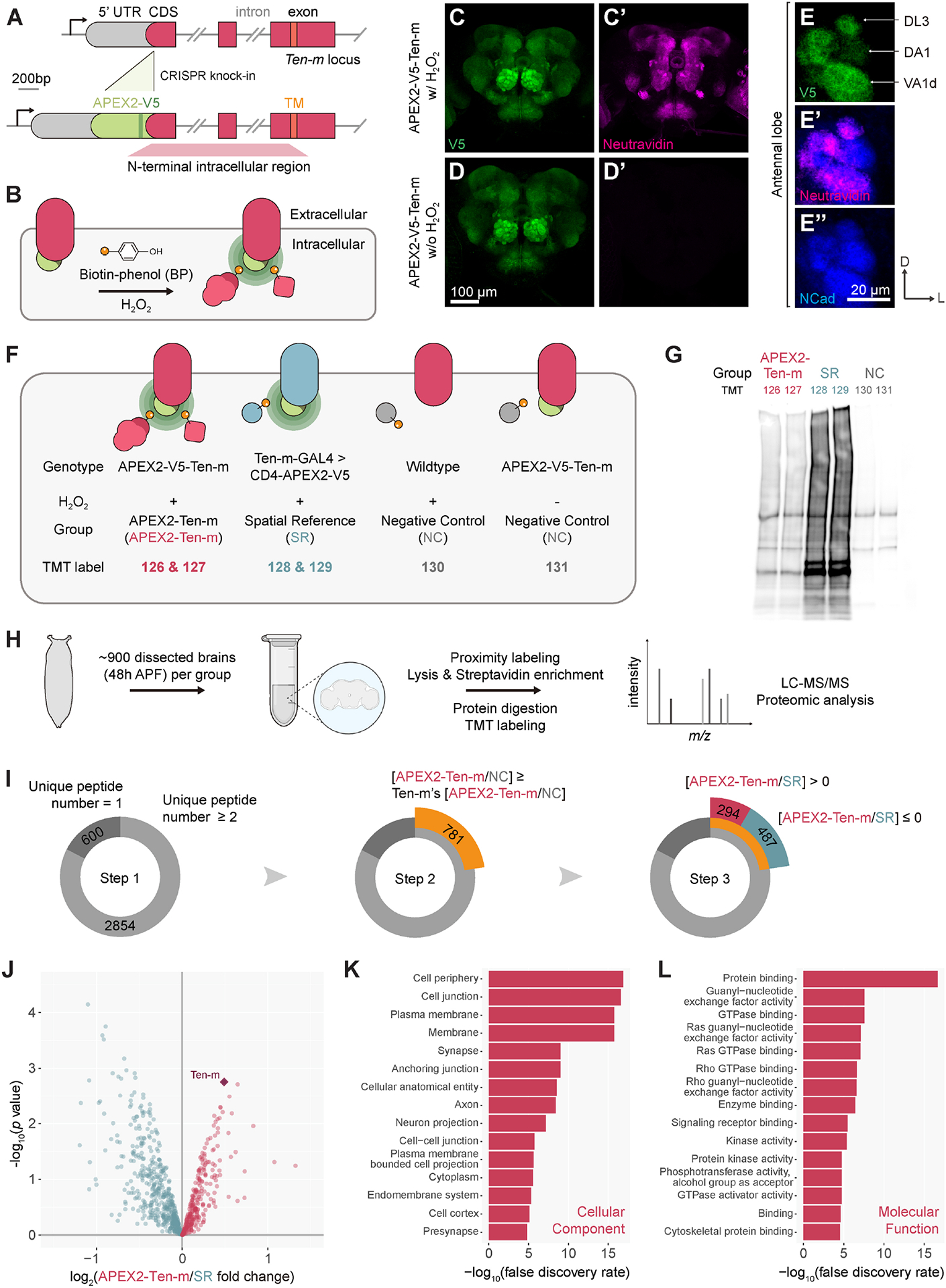
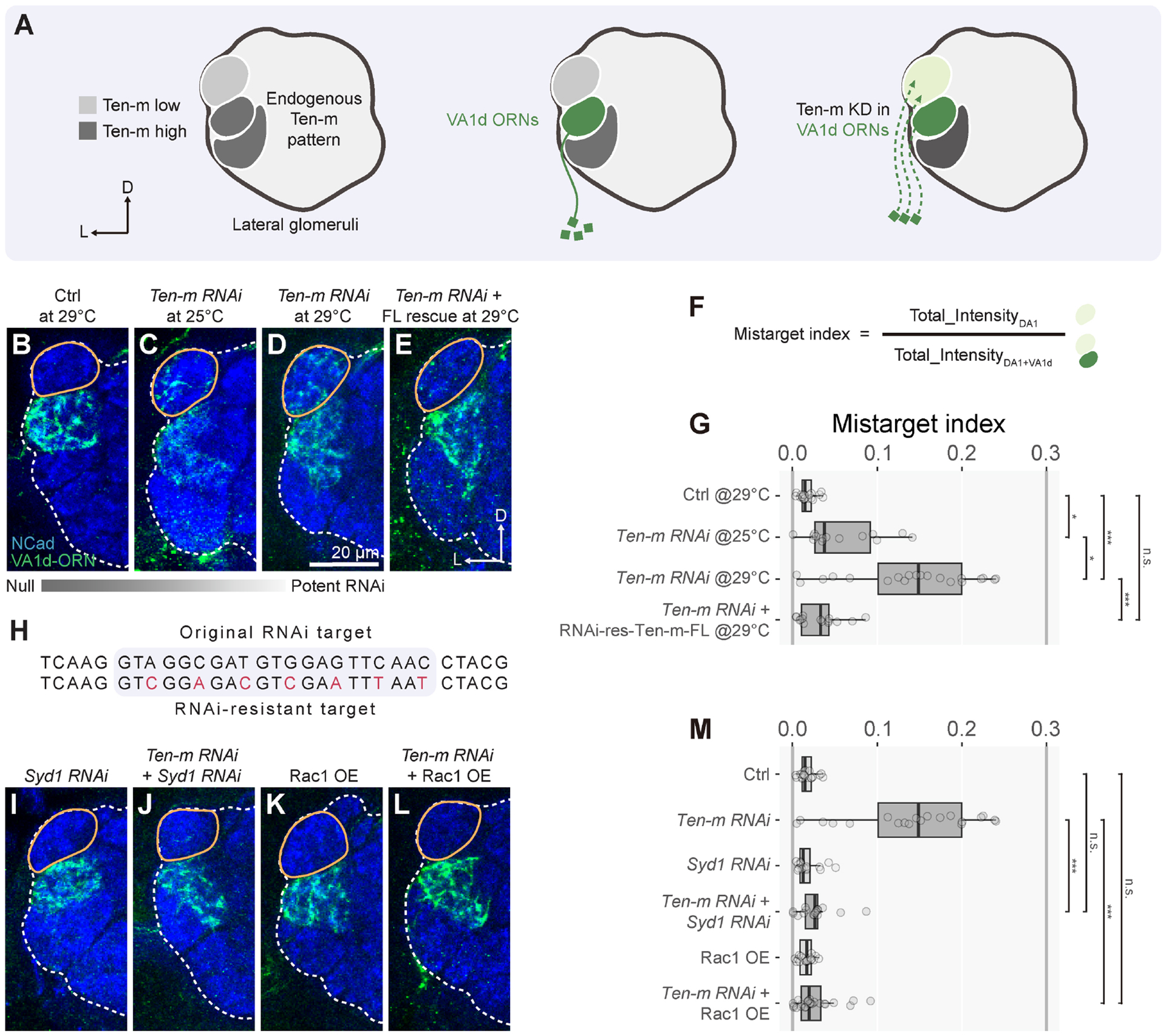
In developing brains, axons exhibit remarkable precision in selecting synaptic partners among many non-partner cells. Evolutionarily conserved teneurins are transmembrane proteins that instruct synaptic partner matching. However, how intracellular signaling pathways execute teneurins' functions is unclear. Here, we use in situ proximity labeling to obtain the intracellular interactome of a teneurin (Ten-m) in the Drosophila brain. Genetic interaction studies using quantitative partner matching assays in both olfactory receptor neurons (ORNs) and projection neurons (PNs) reveal a common pathway: Ten-m binds to and negatively regulates a RhoGAP, thus activating the Rac1 small GTPases to promote synaptic partner matching. Developmental analyses with single-axon resolution identify the cellular mechanism of synaptic partner matching: Ten-m signaling promotes local F-actin levels and stabilizes ORN axon branches that contact partner PN dendrites. Combining spatial proteomics and high-resolution phenotypic analyses, this study advanced our understanding of both cellular and molecular mechanisms of synaptic partner matching.
Keywords: Drosophila; Rho GTPases; RhoGAP; branch stabilization; homophilic attraction; olfactory circuit; olfactory receptor neurons; projection neurons; synaptic partner matching; teneurin.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.L. is a member of the advisory board for Cell.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
