

PTBP1 protects Y RNA from cleavage leading to its apoptosis-specific degradation
- PMID: 38997262
- PMCID: PMC11245482
- DOI: 10.1038/s41420-024-02080-6
PTBP1 protects Y RNA from cleavage leading to its apoptosis-specific degradation
Abstract
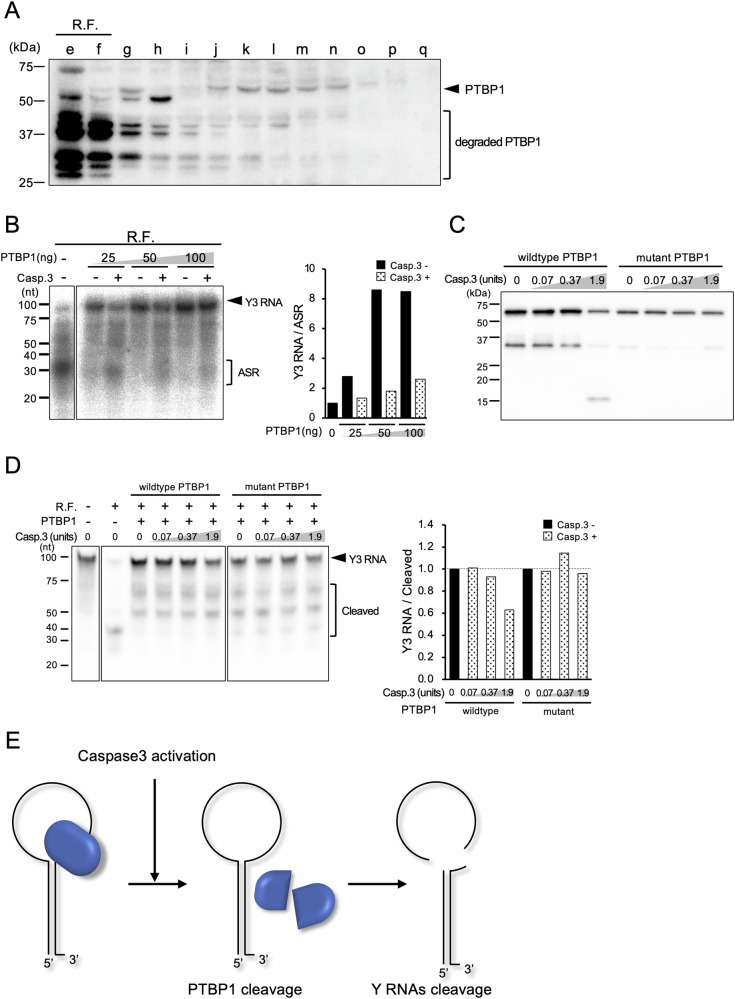
Some RNAs such as 28S rRNA, U1 small nuclear RNA (snRNA), and Y RNAs are known to be cleaved during apoptosis. The underlying mechanism, functions, and biological significance of RNA degradation in apoptosis remain elusive. Y RNAs are non-coding RNAs widely conserved from bacteria to mammals, and are major components of Ro ribonucleoprotein (RNP) complexes which contain the 60 kDa Ro protein (SS-A) and the 50 kDa La protein (SS-B). The autoantigenic Ro and La proteins were identified by autoantibodies present in the sera from patients with Systemic lupus erythematosus (SLE) and Sjögren's syndrome (SjS). We previously identified novel, functional small RNAs named AGO-taxis small RNAs (ASRs) that are specifically bound to Argonaute protein 1 (AGO1), which are processed from Y RNAs. Cell-free analysis combined with fractionation methods revealed that the apoptosis-specific biogenesis of ASRs or cleavage of Y RNA was induced by truncation of polypyrimidine tract-binding protein 1 (PTBP1), which is an endoribonuclease inhibitor of Y RNAs by caspase 3. Caspase 3-resistant PTBP1 mutant protected cleavage of Y RNAs in apoptosis induced by staurosporine. Furthermore, caspase 3-resistant PTBP1 mutant knock-in mice showed elevated cytokines, dysregulation of the germinal center formation compared to the wild-type mice at LPS stimulation, and high positivity of antinuclear antibody. Those results suggest that cleavage of Y RNAs or biogenesis of ASR during apoptosis has critical biological functions and their deregulation result in immune dysregulation and the formation of autoantibody, possibly leading to the development of autoimmune diseases.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Grants and funding
- JP23H02940/MEXT | Japan Society for the Promotion of Science (JSPS)
- 23ama221213h0002/Japan Agency for Medical Research and Development (AMED)
- 23ama221215h0002/Japan Agency for Medical Research and Development (AMED)
- 23fk0210133h0001/Japan Agency for Medical Research and Development (AMED)
- JPMJCR19H5/MEXT | JST | Core Research for Evolutional Science and Technology (CREST)
LinkOut - more resources
Full Text Sources
Research Materials
