

A type II phosphatidylinositol-4-kinase coordinates sorting of cargo polarizing by endocytic recycling
- PMID: 38997419
- PMCID: PMC11245547
- DOI: 10.1038/s42003-024-06553-3
A type II phosphatidylinositol-4-kinase coordinates sorting of cargo polarizing by endocytic recycling
Abstract
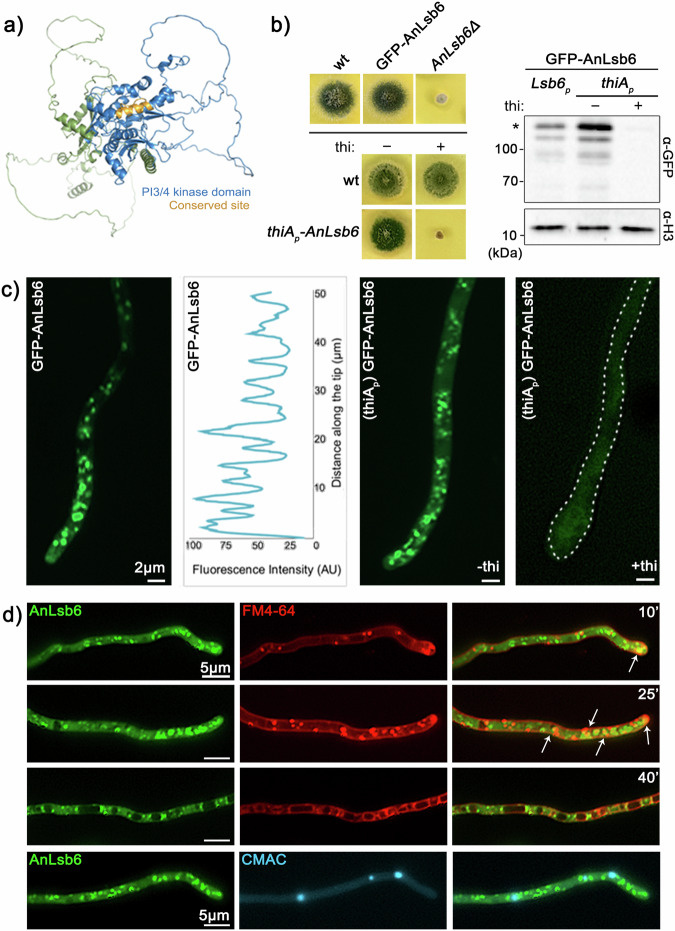
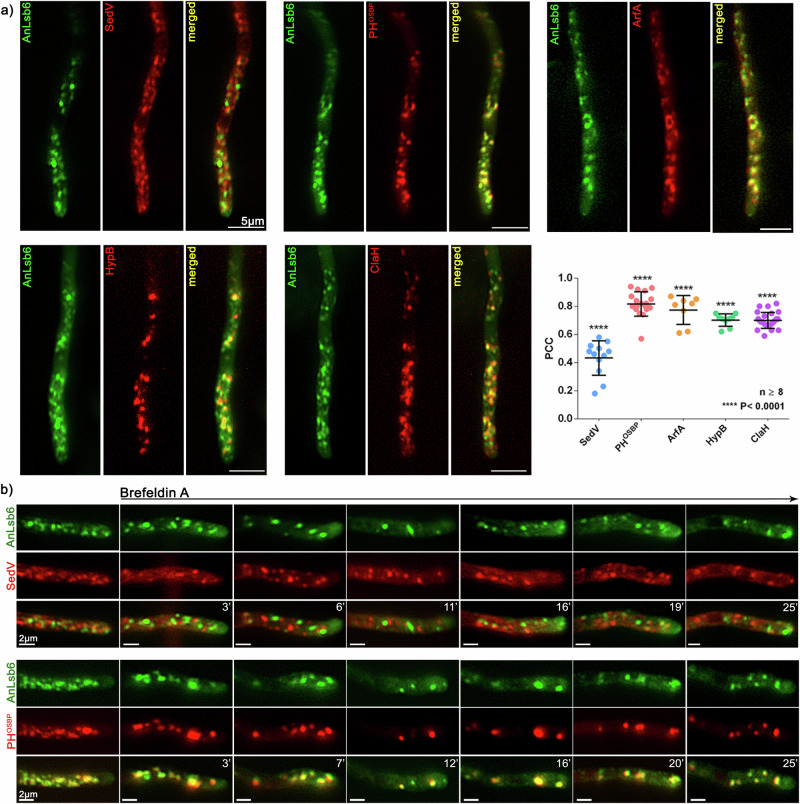
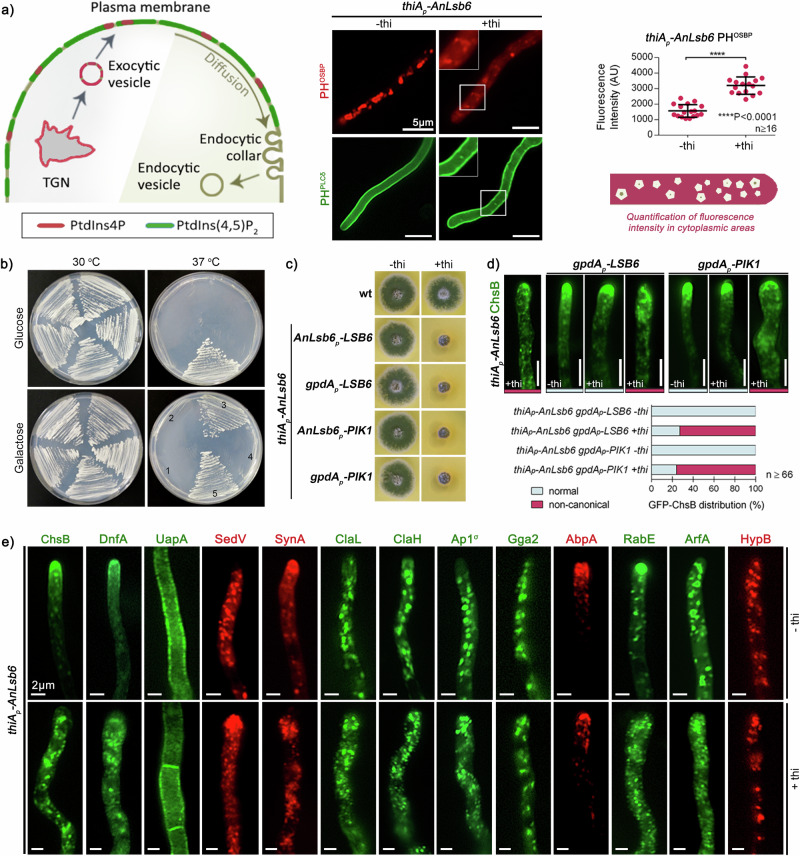
Depending on their phosphorylation status, derivatives of phosphatidylinositol play important roles in vesicle identity, recognition and intracellular trafficking processes. In eukaryotic cells, phosphatidylinositol-4 phosphate pools generated by specific kinases are key determinants of the conventional secretion pathways. Earlier work in yeast has classified phosphatidylinositol-4 kinases in two types, Stt4p and Pik1p belonging to type III and Lsb6p to type II, with distinct cellular localizations and functions. Eurotiomycetes appear to lack Pik1p homologues. In Aspergillus nidulans, unlike homologues in other fungi, AnLsb6 is associated to late Golgi membranes and when heterologously overexpressed, it compensates for the thermosensitive phenotype in a Saccharomyces cerevisiae pik1 mutant, whereas its depletion leads to disorganization of Golgi-associated PHOSBP-labelled membranes, that tend to aggregate dependent on functional Rab5 GTPases. Evidence provided herein, indicates that the single type II phosphatidylinositol-4 kinase AnLsb6 is the main contributor for decorating secretory vesicles with relevant phosphatidylinositol-phosphate species, which navigate essential cargoes following the route of apical polarization via endocytic recycling.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
