

Intermittent Fasting Attenuates Obesity-Induced Triple-Negative Breast Cancer Progression by Disrupting Cell Cycle, Epithelial-Mesenchymal Transition, Immune Contexture, and Proinflammatory Signature
- PMID: 38999849
- PMCID: PMC11243652
- DOI: 10.3390/nu16132101
Intermittent Fasting Attenuates Obesity-Induced Triple-Negative Breast Cancer Progression by Disrupting Cell Cycle, Epithelial-Mesenchymal Transition, Immune Contexture, and Proinflammatory Signature
Abstract
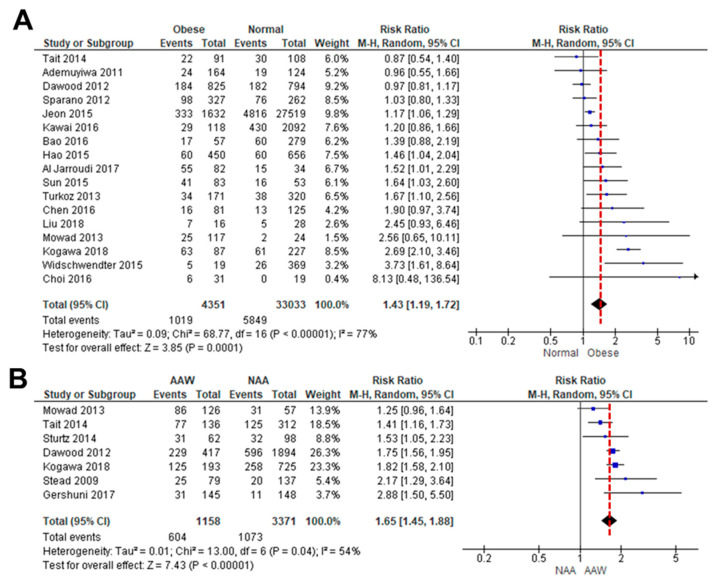
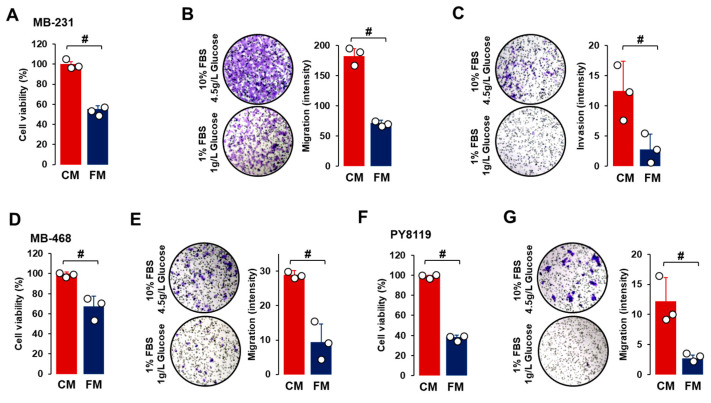
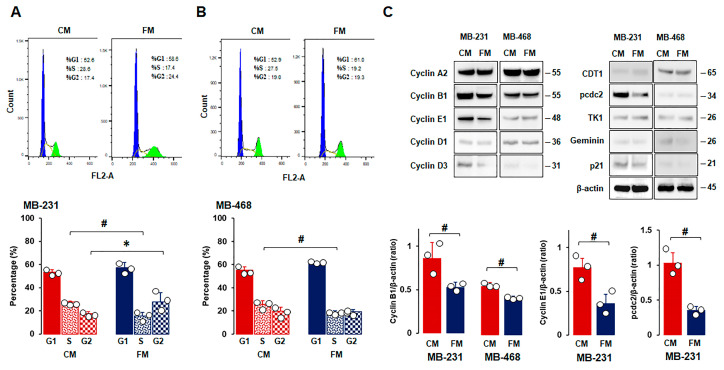
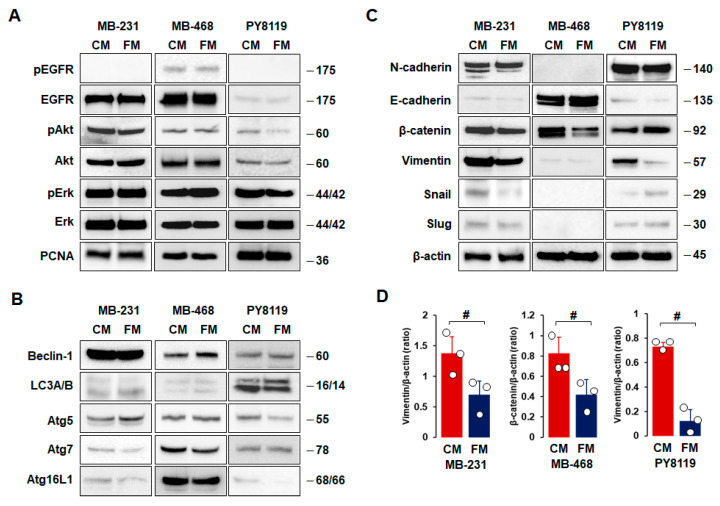
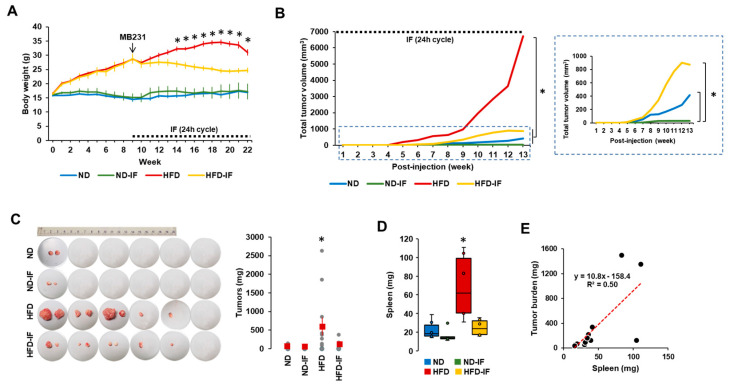
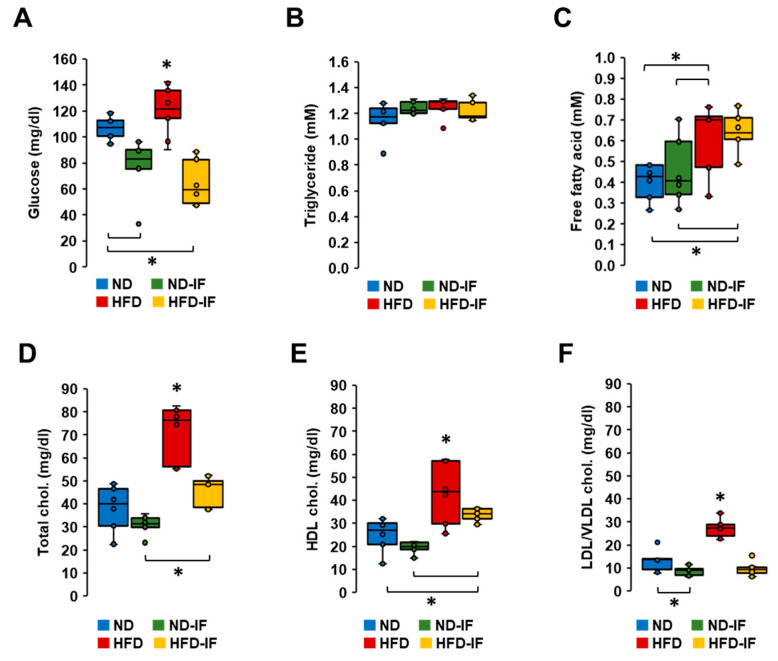
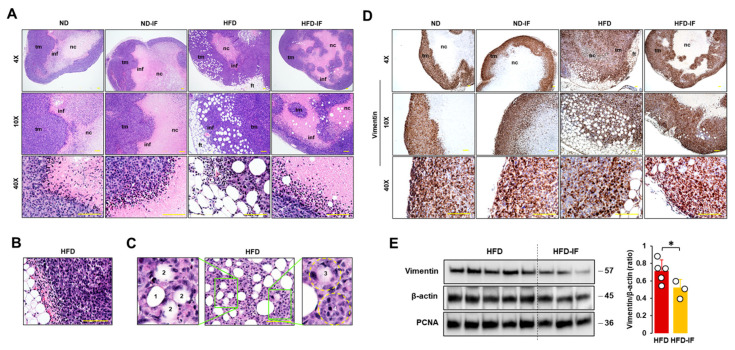
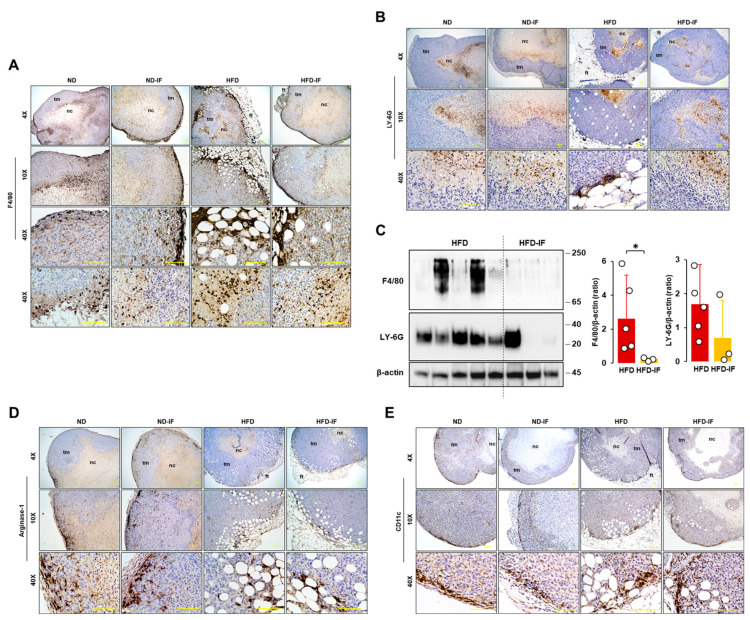
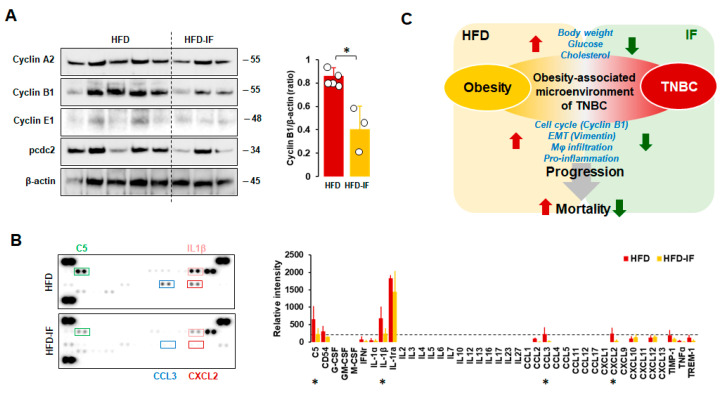
Obesity is associated with one-fifth of cancer deaths, and breast cancer is one of the obesity-related cancers. Triple-negative breast cancer (TNBC) lacks estrogen and progesterone receptors and human epidermal growth factor receptor 2, leading to the absence of these therapeutic targets, followed by poor overall survival. We investigated if obesity could hasten TNBC progression and intermittent fasting (IF) could attenuate the progression of obesity-related TNBC. Our meta-analysis of the TNBC outcomes literature showed that obesity led to poorer overall survival in TNBC patients. Fasting-mimicking media reduced cell proliferation disrupted the cell cycle, and decreased cell migration and invasion. IF decreased body weight in obese mice but no change in normal mice. Obese mice exhibited elevated plasma glucose and cholesterol levels, increased tumor volume and weight, and enhanced macrophage accumulation in tumors. The obesity-exacerbated TNBC progression was attenuated after IF, which decreased cyclin B1 and vimentin levels and reduced the proinflammatory signature in the obesity-associated tumor microenvironment. IF attenuated obesity-induced TNBC progression through reduced obesity and tumor burdens in cell and animal experiments, supporting the potential of a cost-effective adjuvant IF therapy for TNBC through lifestyle change. Further evidence is needed of these IF benefits in TNBC, including from human clinical trials.
Keywords: epithelial–mesenchymal transition; intermittent fasting; obesity; proinflammatory signature; triple-negative breast cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
