

Cortico-basal ganglia plasticity in motor learning
- PMID: 39002543
- PMCID: PMC11309896
- DOI: 10.1016/j.neuron.2024.06.014
Cortico-basal ganglia plasticity in motor learning
Abstract
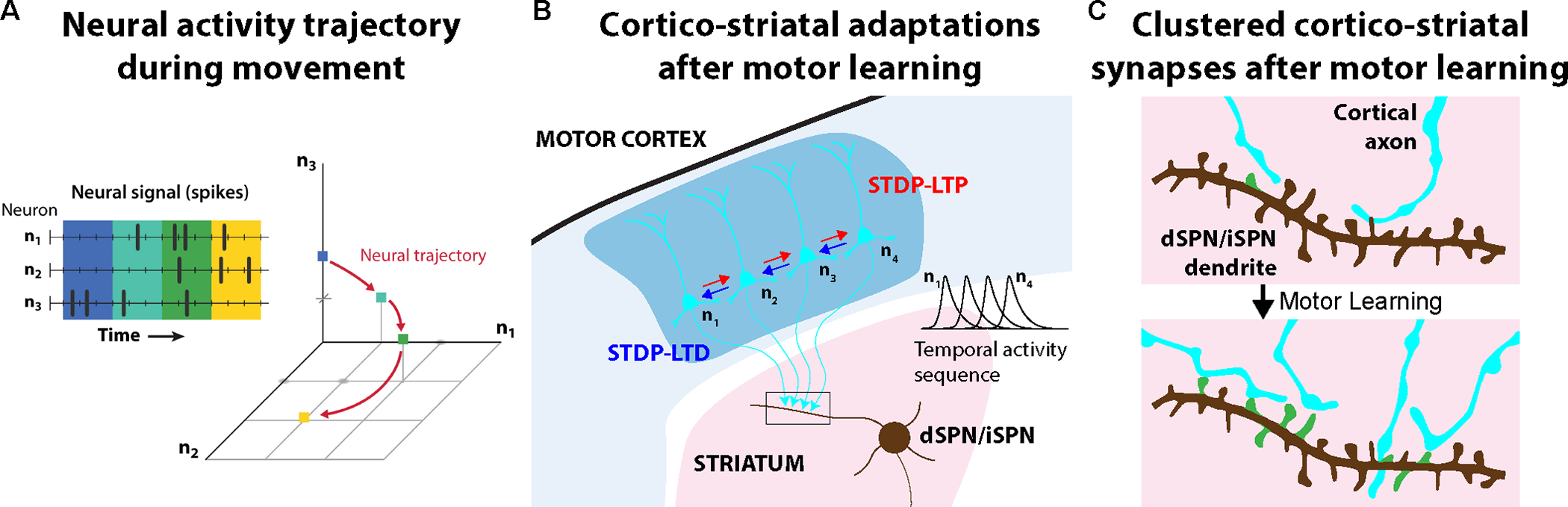
One key function of the brain is to control our body's movements, allowing us to interact with the world around us. Yet, many motor behaviors are not innate but require learning through repeated practice. Among the brain's motor regions, the cortico-basal ganglia circuit is particularly crucial for acquiring and executing motor skills, and neuronal activity in these regions is directly linked to movement parameters. Cell-type-specific adaptations of activity patterns and synaptic connectivity support the learning of new motor skills. Functionally, neuronal activity sequences become structured and associated with learned movements. On the synaptic level, specific connections become potentiated during learning through mechanisms such as long-term synaptic plasticity and dendritic spine dynamics, which are thought to mediate functional circuit plasticity. These synaptic and circuit adaptations within the cortico-basal ganglia circuitry are thus critical for motor skill acquisition, and disruptions in this plasticity can contribute to movement disorders.
Keywords: basal ganglia; circuit adaptation; dendritic spines; motor control; motor cortex; motor learning; striatum; synaptic plasticity.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
