

Redundancy in microbiota-mediated suppression of the soybean cyst nematode
- PMID: 39004755
- PMCID: PMC11247744
- DOI: 10.1186/s40168-024-01840-x
Redundancy in microbiota-mediated suppression of the soybean cyst nematode
Abstract
Background: Soybean cyst nematodes (SCN) as animal parasites of plants are not usually interested in killing the host but are rather focused on completing their life cycle to increase population, resulting in substantial yield losses. Remarkably, some agricultural soils after long-term crop monoculture show a significant decline in SCN densities and suppress disease in a sustainable and viable manner. However, relatively little is known about the microbes and mechanisms operating against SCN in such disease-suppressive soils.
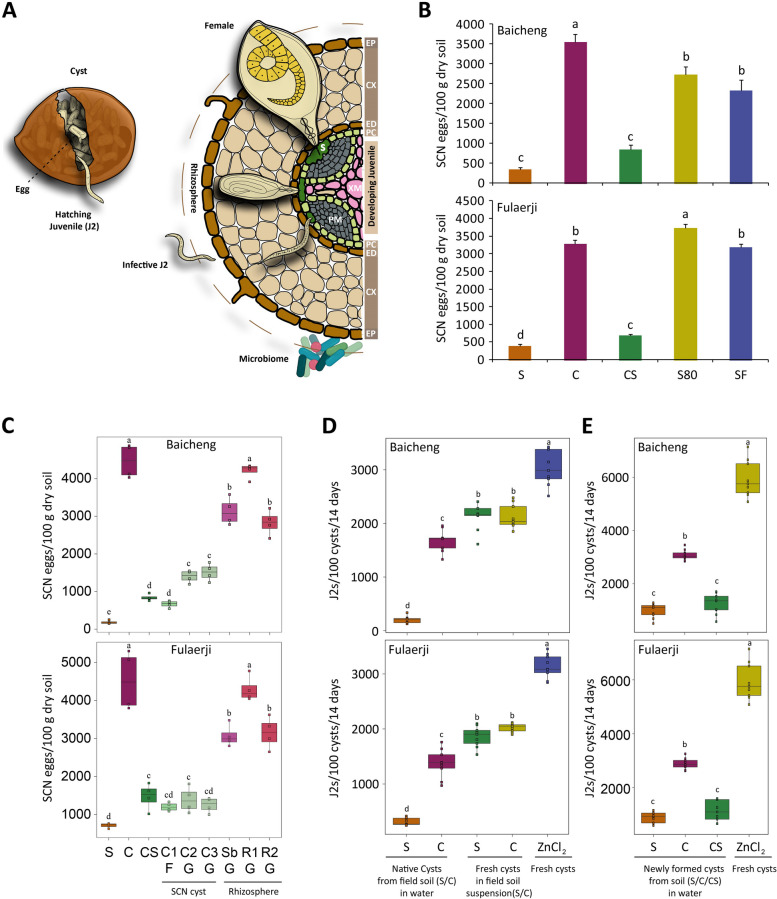
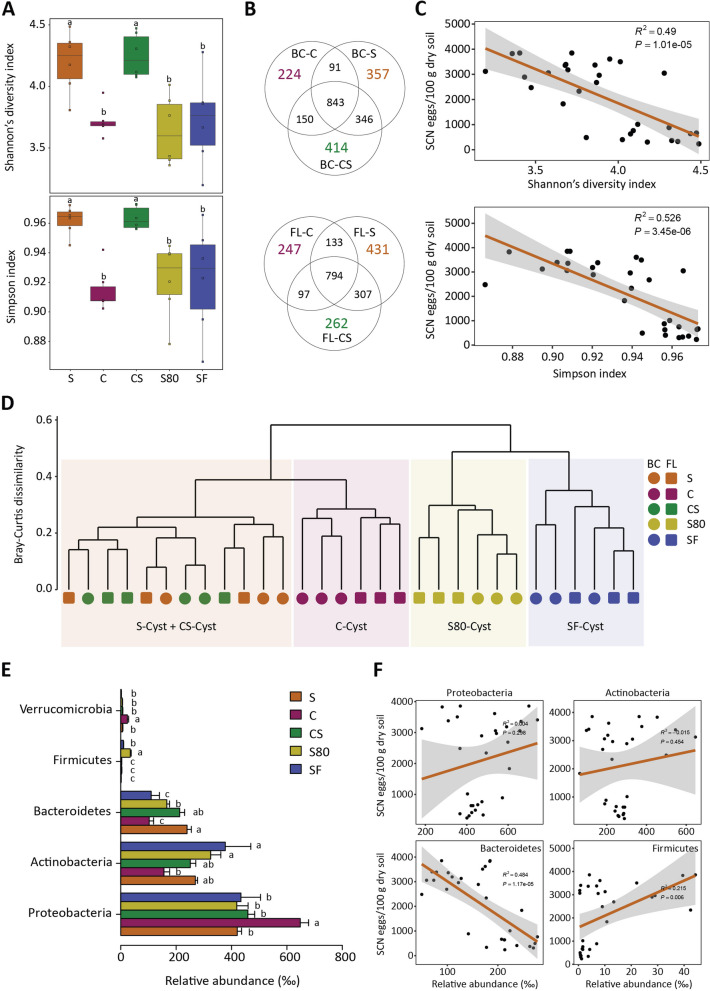
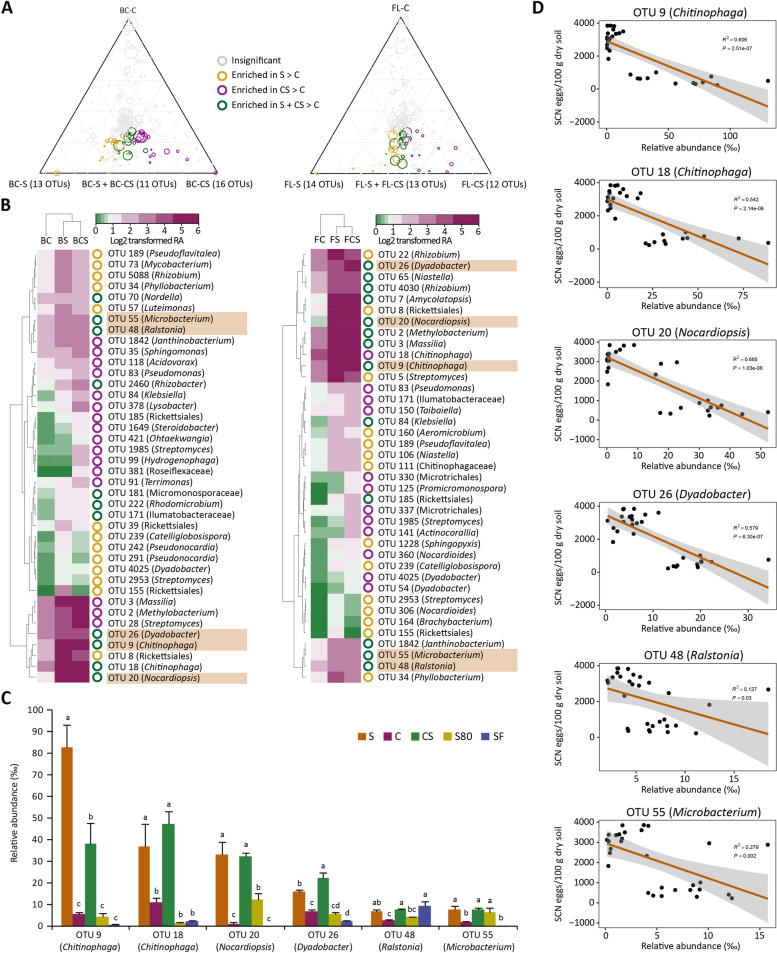
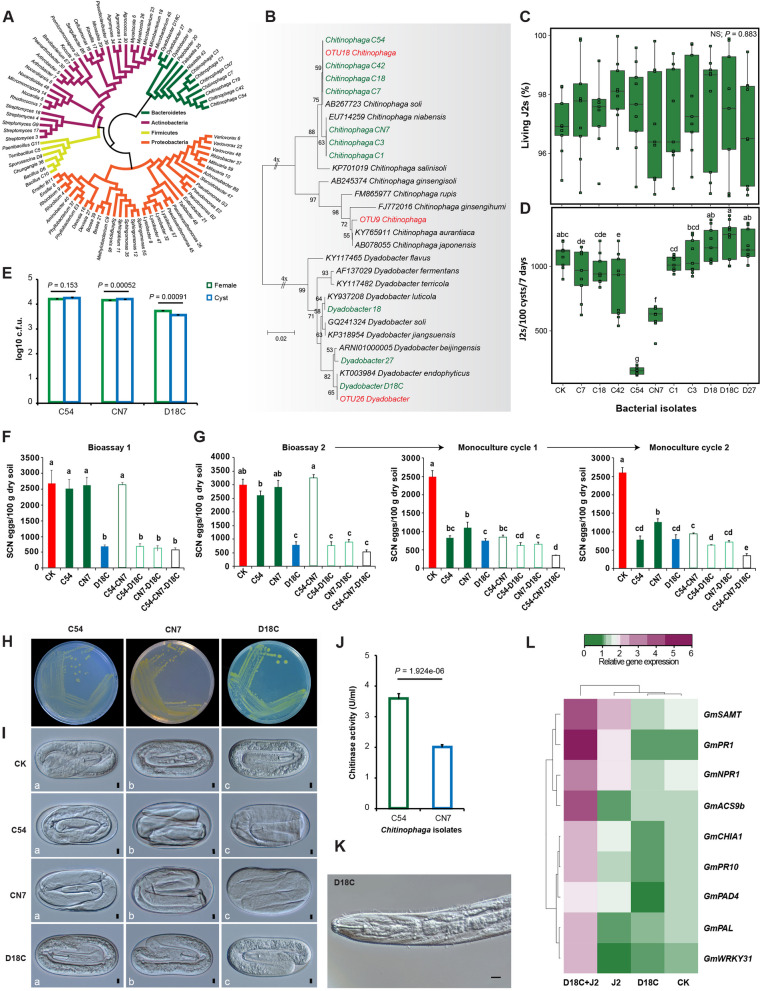
Results: Greenhouse experiments showed that suppressive soils (S) collected from two provinces of China and transplantation soils (CS, created by mixing 10% S with 90% conducive soils) suppressed SCN. However, SCN suppressiveness was partially lost or completely abolished when S soils were treated with heat (80 °C) and formalin. Bacterial community analysis revealed that the specific suppression in S and CS was mainly associated with the bacterial phylum Bacteroidetes, specifically due to the enrichment of Chitinophaga spp. and Dyadobacter sp., in the cysts. SCN cysts colonized by Chitinophaga spp. showed dramatically reduced egg hatching, with unrecognizable internal body organization of juveniles inside the eggshell due to chitinase activity. Whereas, Dyadobacter sp. cells attached to the surface coat of J2s increased soybean resistance against SCN by triggering the expression of defence-associated genes. The disease-suppressive potential of these bacteria was validated by inoculating them into conducive soil. The Dyadobacter strain alone or in combination with Chitinophaga strains significantly decreased egg densities after one growing cycle of soybeans. In contrast, Chitinophaga strains alone required more than one growing cycle to significantly reduce SCN egg hatching and population density.
Conclusion: This study revealed how soybean monoculture for decades induced microbiota homeostasis, leading to the formation of SCN-suppressive soil. The high relative abundance of antagonistic bacteria in the cyst suppressed the SCN population both directly and indirectly. Because uncontrolled proliferation will likely lead to quick demise due to host population collapse, obligate parasites like SCN may have evolved to modulate virulence/proliferation to balance these conflicting needs. Video Abstract.
Keywords: Heterodera glycines; Chitinase; Cyst microbiota; Induced resistance; Nematode suppression; Soybean; Suppressive soil.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Liu XB, Herbert SJ. Fifteen years of research examining cultivation of continuous soybean in northeast China: a review. Field Crops Res. 2002;79:1–7. doi: 10.1016/S0378-4290(02)00042-4. - DOI
MeSH terms
Substances
Grants and funding
- PEN4839/USDA National Institute of Food & Agriculture and Hatch Appropriations
- 2022YFD1400700/National Key Research and Development Program of China
- 2022YFD1400700/National Key Research and Development Program of China
- 31770065/National Natural Science Foundation of China
- 31770065/National Natural Science Foundation of China
LinkOut - more resources
Full Text Sources
Miscellaneous
