

This is a preprint.
Postprandial sleep in short-sleeping Mexican cavefish
- PMID: 39005273
- PMCID: PMC11244998
- DOI: 10.1101/2024.07.03.602003
Postprandial sleep in short-sleeping Mexican cavefish
Update in
-
Postprandial Sleep in Short-Sleeping Mexican Cavefish.J Exp Zool A Ecol Integr Physiol. 2024 Dec;341(10):1084-1096. doi: 10.1002/jez.2880. Epub 2024 Nov 13. J Exp Zool A Ecol Integr Physiol. 2024. PMID: 39539086 Free PMC article.
Abstract
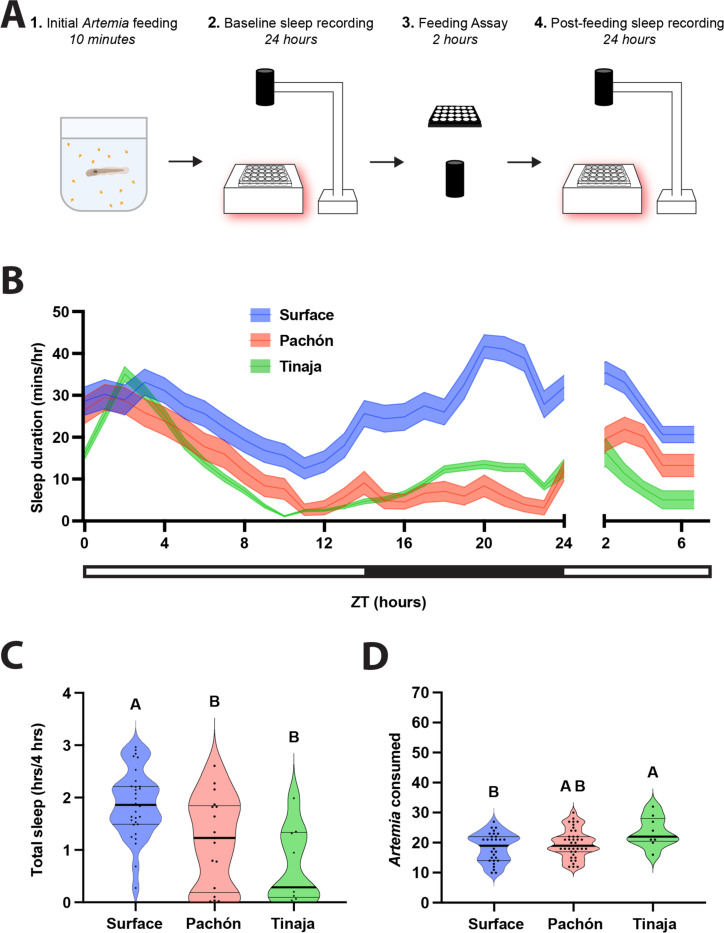
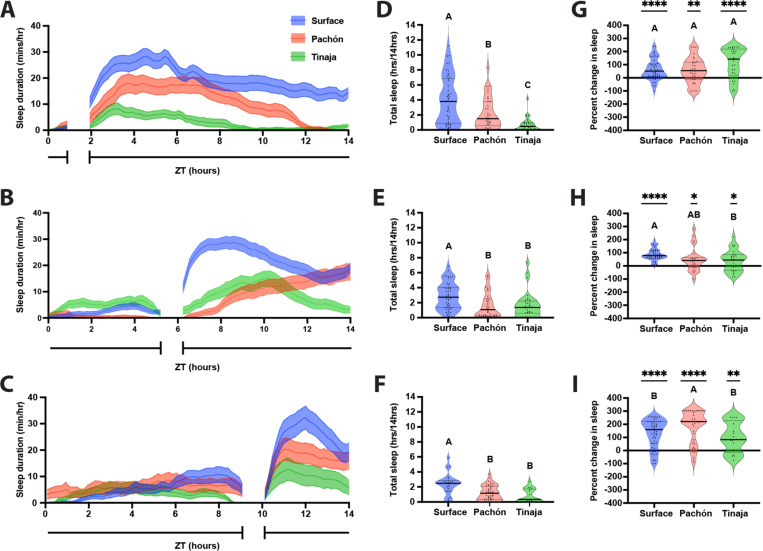
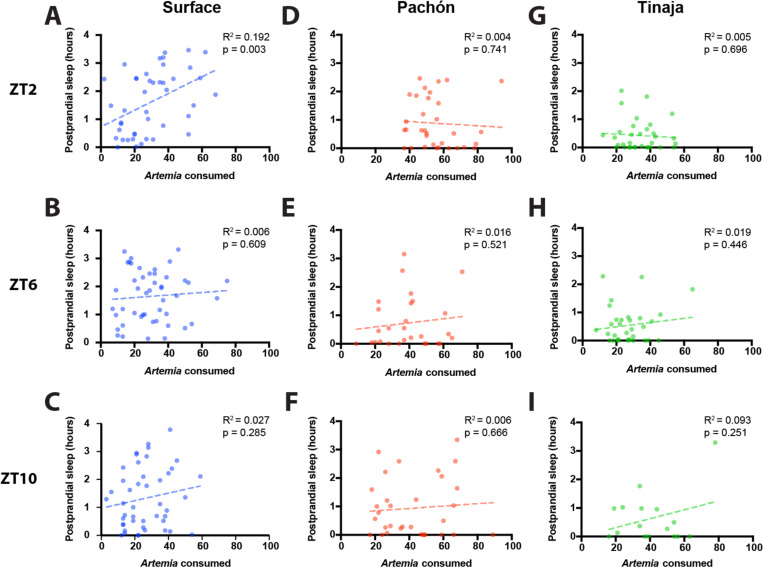
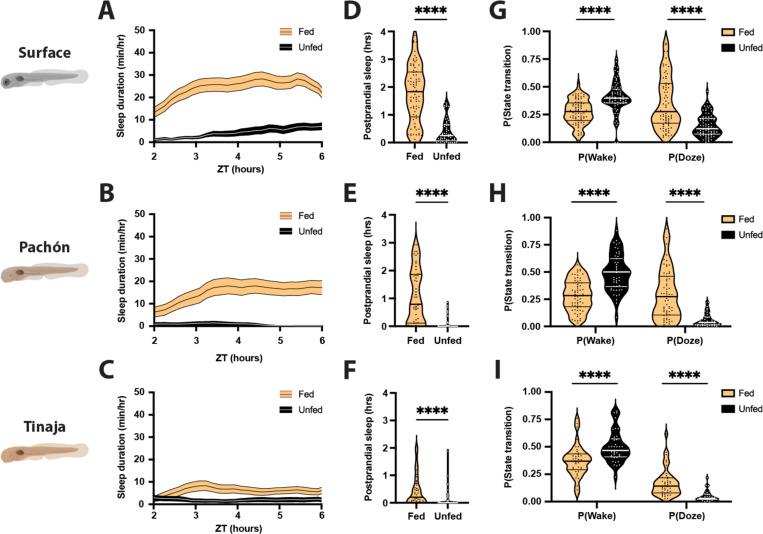
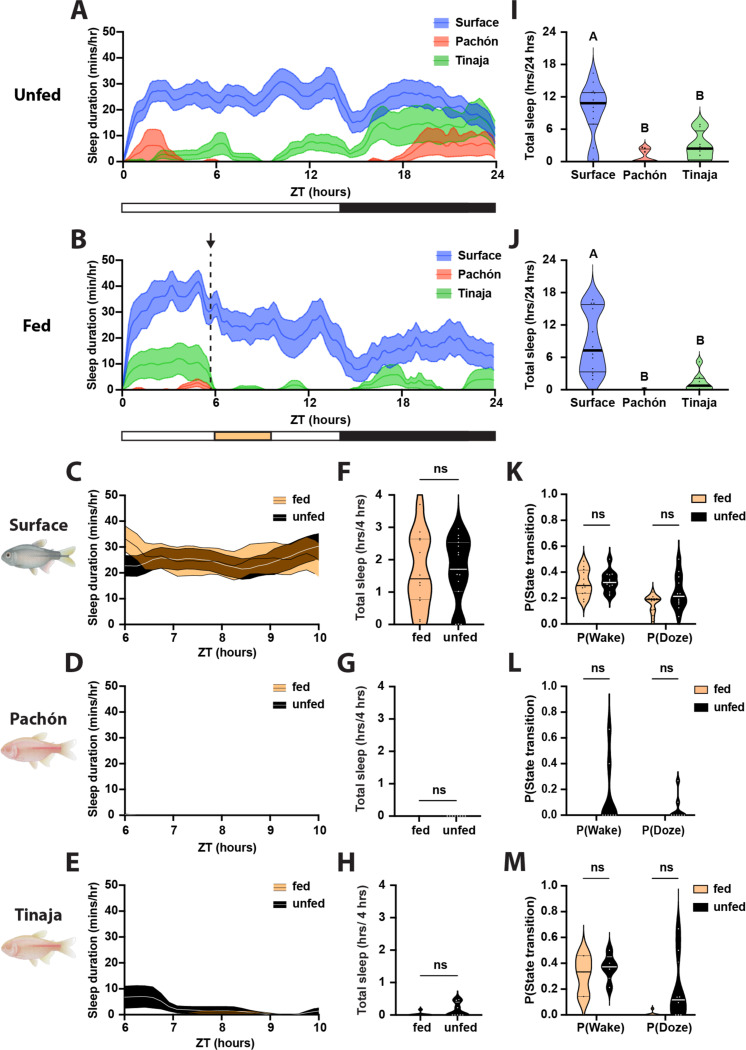
Interaction between sleep and feeding behaviors are critical for adaptive fitness. Diverse species suppress sleep when food is scarce to increase the time spent foraging. Post-prandial sleep, an increase in sleep time following a feeding event, has been documented in vertebrate and invertebrate animals. While interactions between sleep and feeding appear to be highly conserved, the evolution of postprandial sleep in response to changes in food availability remains poorly understood. Multiple populations of the Mexican cavefish, Astyanax mexicanus, have independently evolved sleep loss and increased food consumption compared to surface-dwelling fish of the same species, providing the opportunity to investigate the evolution of interactions between sleep and feeding. Here, we investigate effects of feeding on sleep in larval and adult surface fish, and two parallelly evolved cave populations of A. mexicanus. Larval surface and cave populations of A. mexicanus increase sleep immediately following a meal, providing the first evidence of postprandial sleep in a fish model. The amount of sleep was not correlated to meal size and occurred independently of feeding time. In contrast to larvae, postprandial sleep was not detected in adult surface or cavefish, that can survive for months without food. Together, these findings reveal that postprandial sleep is present in multiple short-sleeping populations of cavefish, suggesting sleep-feeding interactions are retained despite the evolution of sleep loss. These findings raise the possibility that postprandial sleep is critical for energy conservation and survival in larvae that are highly sensitive to food deprivation.
Figures
References
-
- Anafi R. C., Kayser M. S., and Raizen D. M., 2019. Exploring phylogeny to find the function of sleep. Nat Rev Neurosci 20: 109–116. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources