

This is a preprint.
Frequent transitions in self-assembly across the evolution of a central metabolic enzyme
- PMID: 39005358
- PMCID: PMC11245102
- DOI: 10.1101/2024.07.05.602260
Frequent transitions in self-assembly across the evolution of a central metabolic enzyme
Update in
-
Frequent transitions in self-assembly across the evolution of a central metabolic enzyme.Nat Commun. 2024 Dec 3;15(1):10515. doi: 10.1038/s41467-024-54408-6. Nat Commun. 2024. PMID: 39627196 Free PMC article.
Abstract
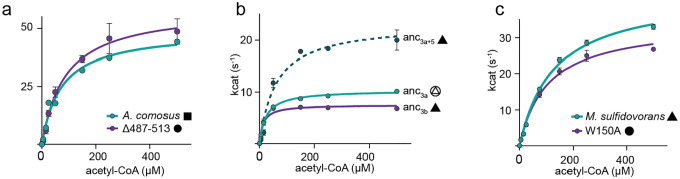
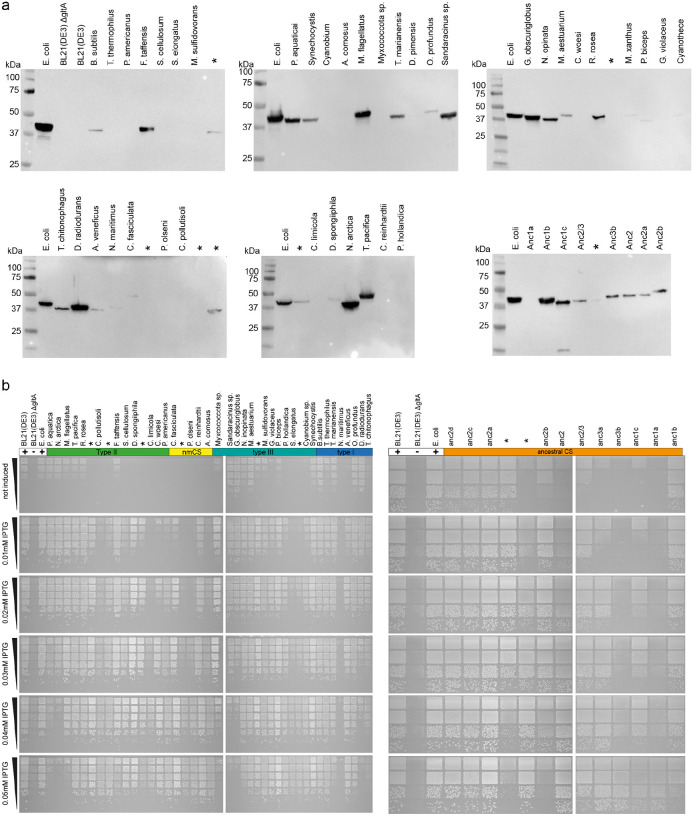
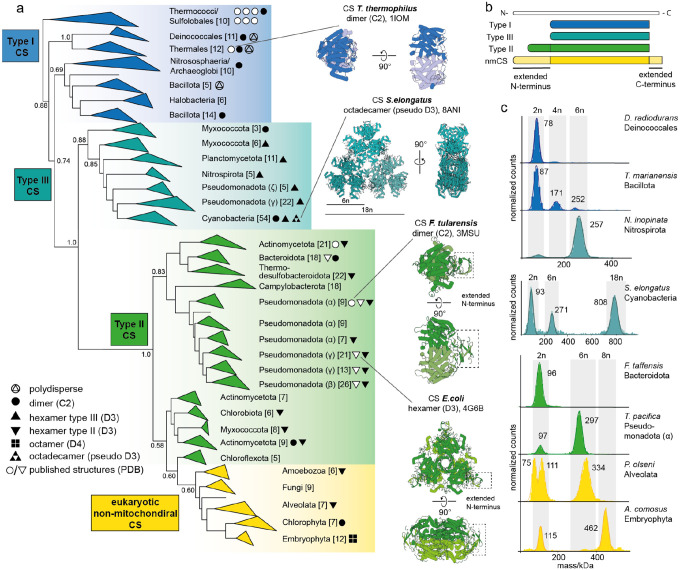
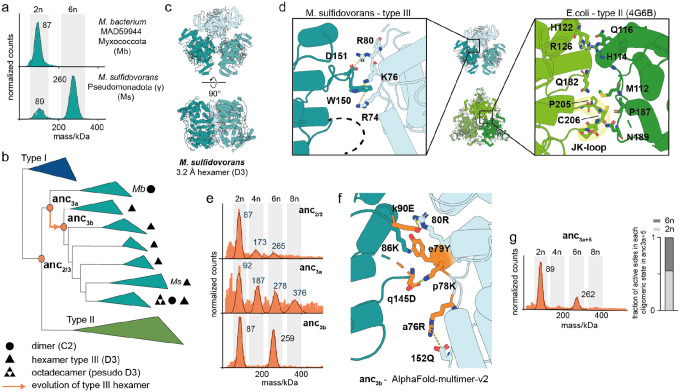
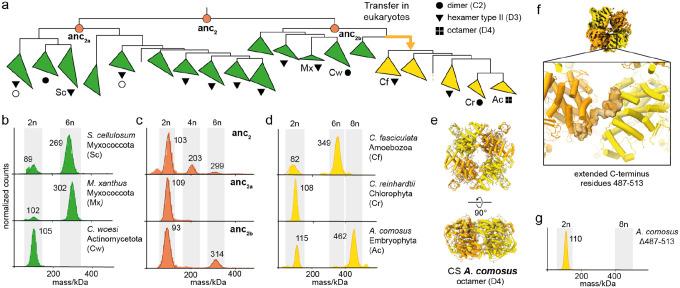
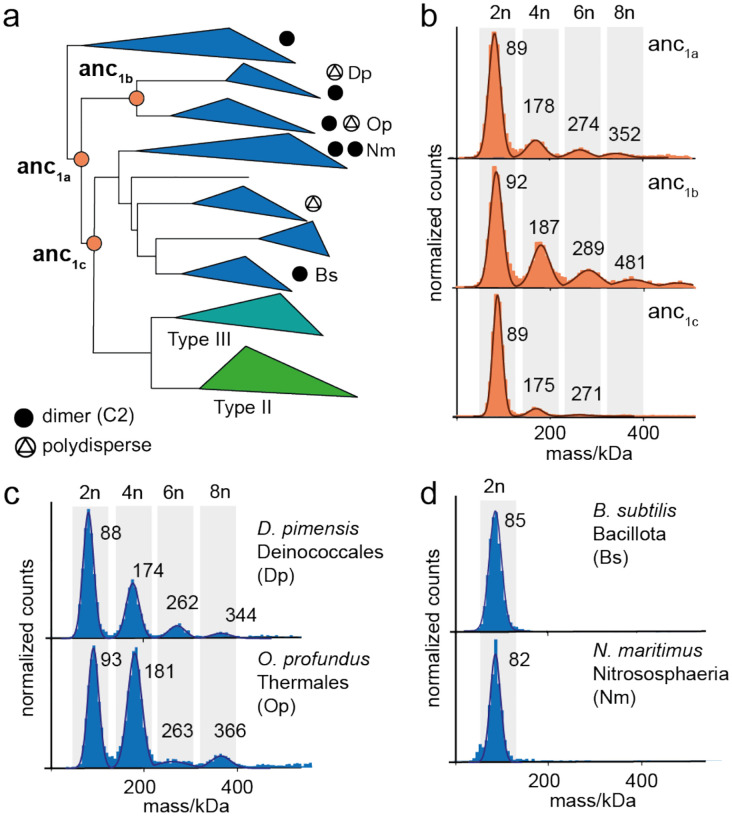
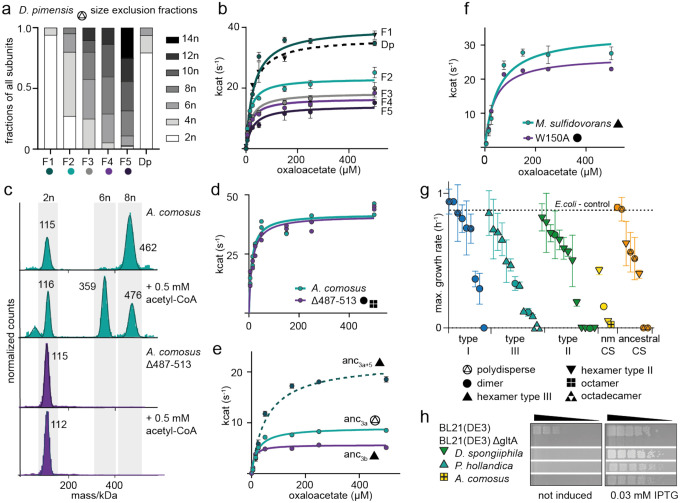
Many enzymes assemble into homomeric protein complexes comprising multiple copies of one protein. Because structural form is usually assumed to follow function in biochemistry, these assemblies are thought to evolve because they provide some functional advantage. In many cases, however, no specific advantage is known and, in some cases, quaternary structure varies among orthologs. This has led to the proposition that self-assembly may instead vary neutrally within protein families. The extent of such variation has been difficult to ascertain because quaternary structure has until recently been difficult to measure on large scales. Here, we employ mass photometry, phylogenetics, and structural biology to interrogate the evolution of homo-oligomeric assembly across the entire phylogeny of prokaryotic citrate synthases - an enzyme with a highly conserved function. We discover a menagerie of different assembly types that come and go over the course of evolution, including cases of parallel evolution and reversions from complex to simple assemblies. Functional experiments in vitro and in vivo indicate that evolutionary transitions between different assemblies do not strongly influence enzyme catalysis. Our work suggests that enzymes can wander relatively freely through a large space of possible assemblies and demonstrates the power of characterizing structure-function relationships across entire phylogenies.
Conflict of interest statement
Competing interests: The authors declare no other competing interests.
Figures

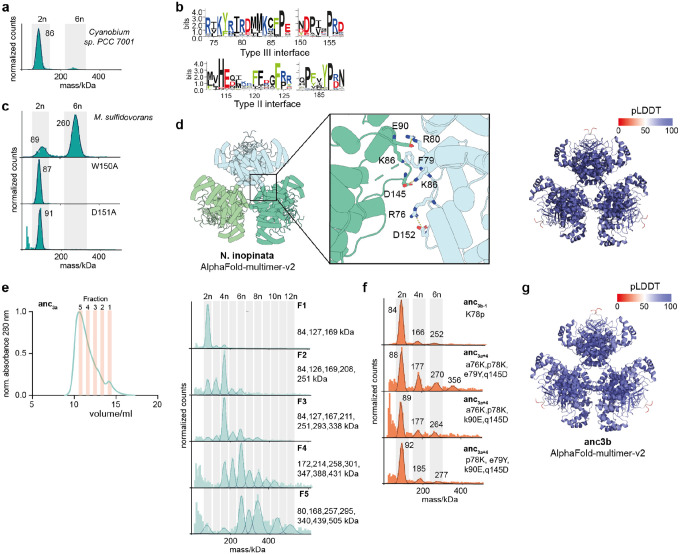
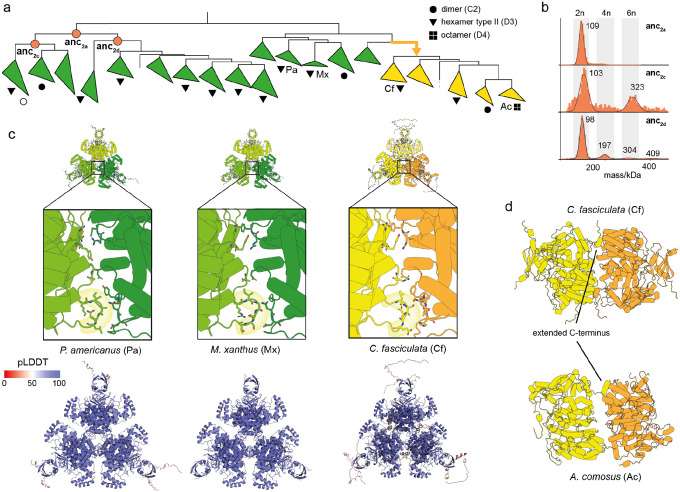
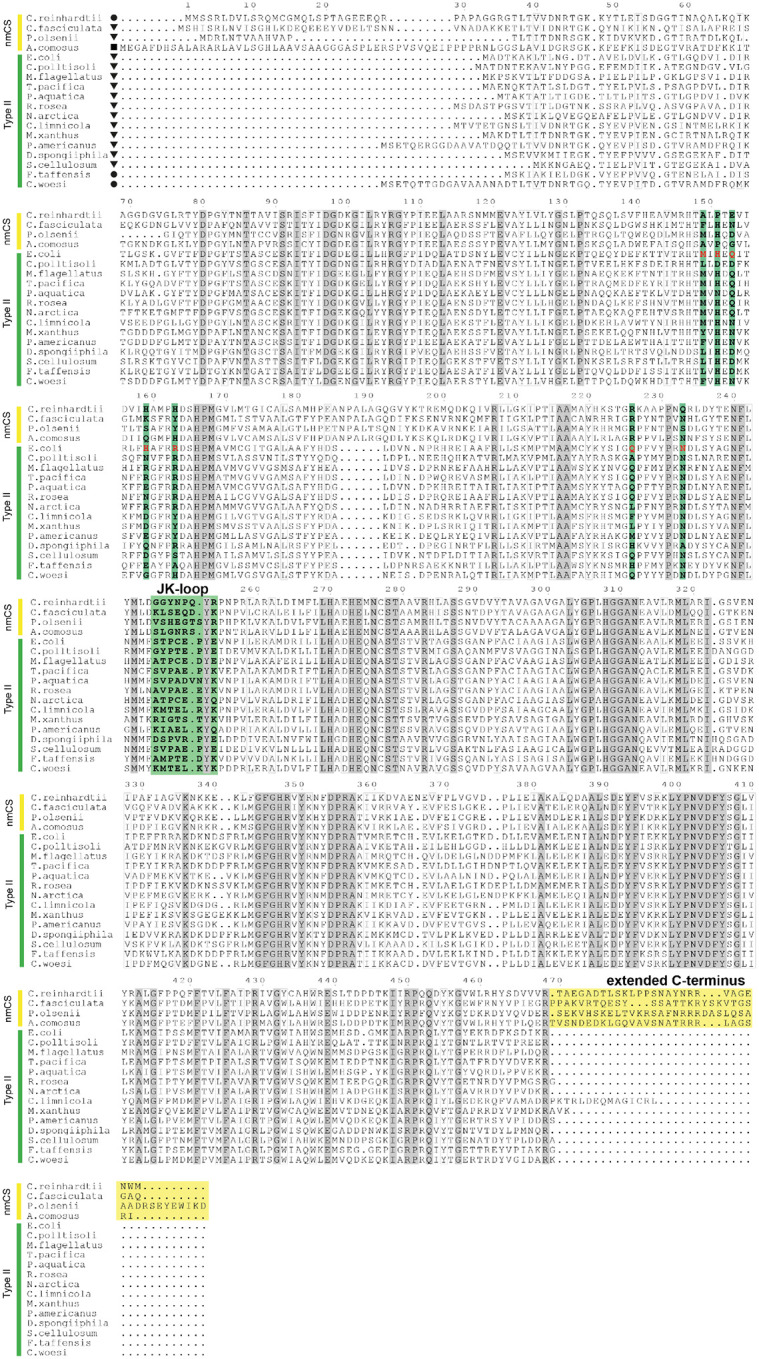
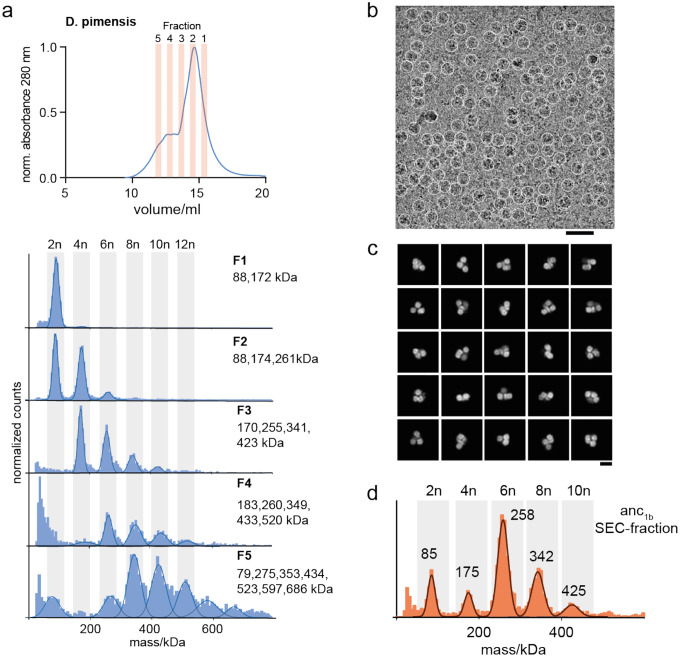
References
-
- Goodsell D. S. & Olson A. J. Structural symmetry and protein function. Annu. Rev. Biophys. Biomol. Struct. 29, 105–153 (2000). - PubMed
-
- Levy E. D. & Teichmann S. Structural, evolutionary, and assembly principles of protein oligomerization. in Progress in Molecular Biology and Translational Science vol. 117 25–51 (Elsevier B.V., 2013). - PubMed
-
- Perica T. et al. The emergence of protein complexes: quaternary structure, dynamics and allostery. Biochem. Soc. Trans. 40, 475–491 (2012). - PubMed
-
- Marianayagam NJ, Sunde M, M. J. The power of two: protein dimerization in biology. Trends Biochem. Sci. 29, 618–625 (2004). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources