

This is a preprint.
A genome-wide association study implicates the olfactory system in Drosophila diapause-associated lifespan extension and fecundity
- PMID: 39005458
- PMCID: PMC11244867
- DOI: 10.1101/2024.03.10.584341
A genome-wide association study implicates the olfactory system in Drosophila diapause-associated lifespan extension and fecundity
Update in
-
A genome-wide association study implicates the olfactory system in Drosophila melanogaster diapause-associated lifespan extension and fecundity.Elife. 2025 Jul 22;13:RP98142. doi: 10.7554/eLife.98142. Elife. 2025. PMID: 40693912 Free PMC article.
Abstract
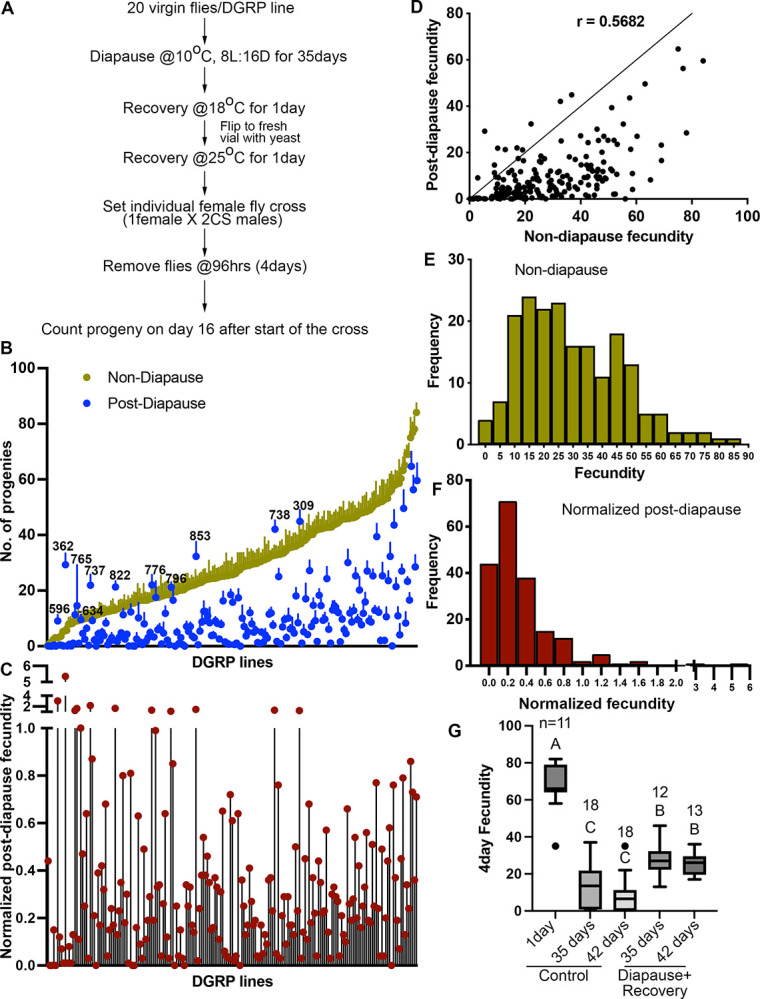
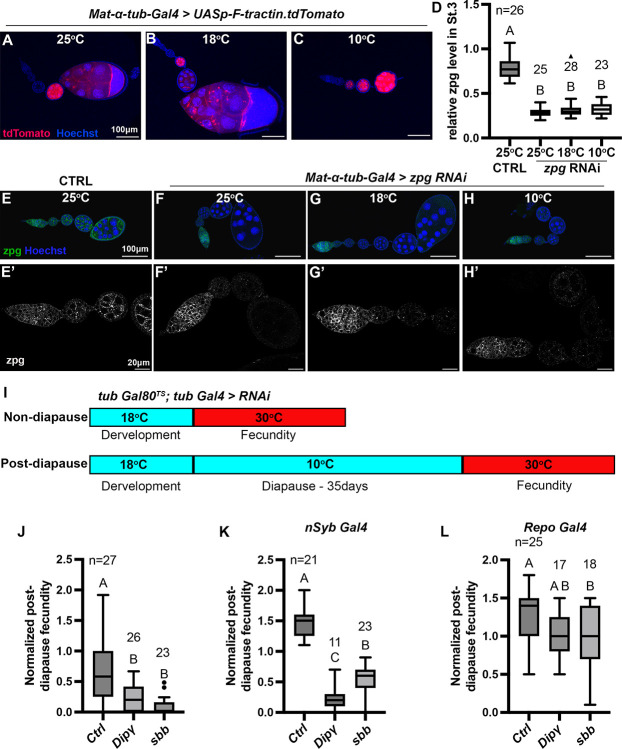
The effects of environmental stress on animal life are gaining importance. Diapause is a reversible dormancy program triggered by adverse environmental conditions. To characterize the genetic basis of this complex program, we leveraged the Drosophila Genome Reference Panel (DGRP) to conduct a Genome-Wide Association Study (GWAS). We assessed post-diapause and non-diapause fecundity across 193 DGRP lines. GWAS revealed 546 variants, encompassing single nucleotide polymorphisms, insertions, and deletions associated with post-diapause fecundity. We identified 291 candidate diapause-associated genes, 40 of which were previously associated with diapause. Gene network analysis indicated that the diapause-associated genes were primarily linked to neuronal and reproductive system development. Similarly, comparison with other fly GWAS revealed the greatest overlap with olfactory-behavior-associated and fecundity-and-lifespan-associated genes. An RNAi screen of selected candidates identified two neuronal genes, Dip-γ and Scribbler, to be required during recovery for post-diapause fecundity. To complement the genetic analysis, we tested which neurons are required for successful diapause. Although amputation of the antenna had little effect on non-diapause lifespan, it reduced diapause lifespan and post-diapause fecundity. Furthermore, olfactory receptor neurons and temperature-sensing neurons were required for successful diapause recovery. Our results provide insights into the molecular, cellular, and genetic basis of Drosophila adult reproductive diapause.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures



References
-
- Budelli G, Ni L, Berciu C, van Giesen L, Knecht ZA, Chang EC, Kaminski B, Silbering AF, Samuel A, Klein M, Benton R, Nicastro D, Garrity PA. 2019. Ionotropic receptors specify the morphogenesis of phasic sensors controlling rapid thermal preference in drosophila. Neuron 101:738–747.e3. doi: 10.1016/j.neuron.2018.12.022 - DOI - PMC - PubMed
-
- Clarke DJB, Jeon M, Stein DJ, Moiseyev N, Kropiwnicki E, Dai C, Xie Z, Wojciechowicz ML, Litz S, Hom J, Evangelista JE, Goldman L, Zhang S, Yoon C, Ahamed T, Bhuiyan S, Cheng M, Karam J, Jagodnik KM, Shu I, Ma’ayan A. 2021. Appyters: Turning Jupyter Notebooks into data-driven web apps. Patterns (N Y) 2:100213. doi: 10.1016/j.patter.2021.100213 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources