

Adenovirus small E1A directs activation of Alu transcription at YAP/TEAD- and AP-1-bound enhancers through interactions with the EP400 chromatin remodeler
- PMID: 39011896
- PMCID: PMC11381368
- DOI: 10.1093/nar/gkae615
Adenovirus small E1A directs activation of Alu transcription at YAP/TEAD- and AP-1-bound enhancers through interactions with the EP400 chromatin remodeler
Abstract
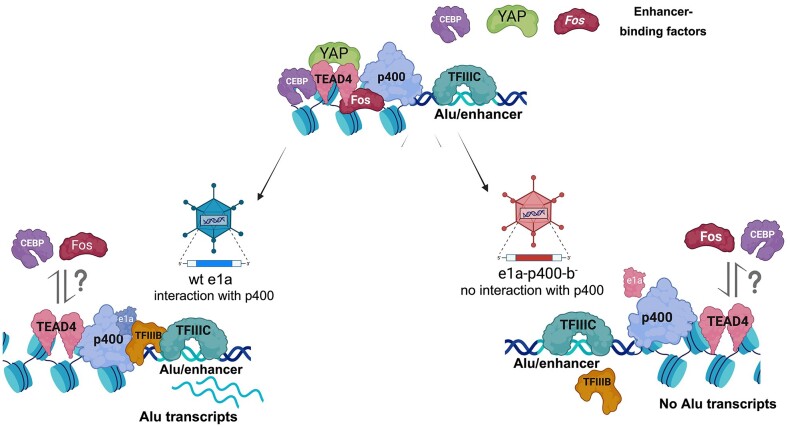
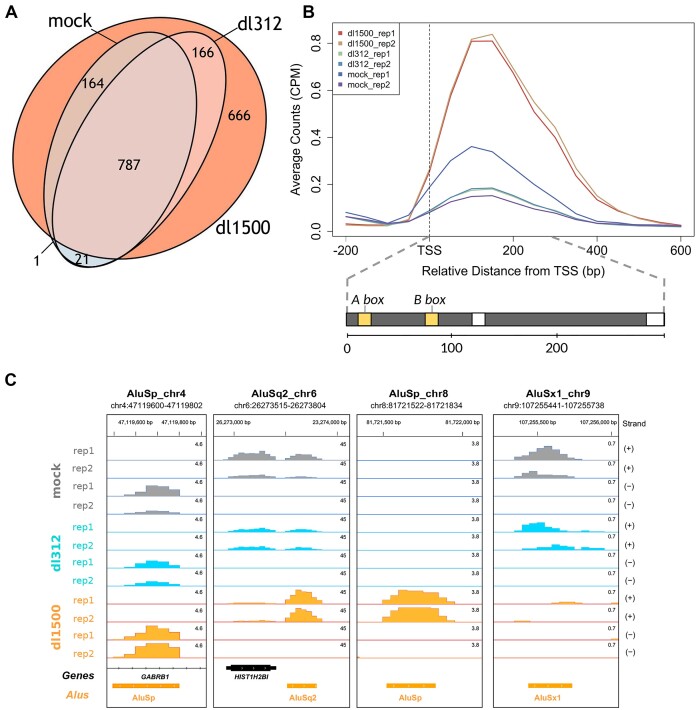
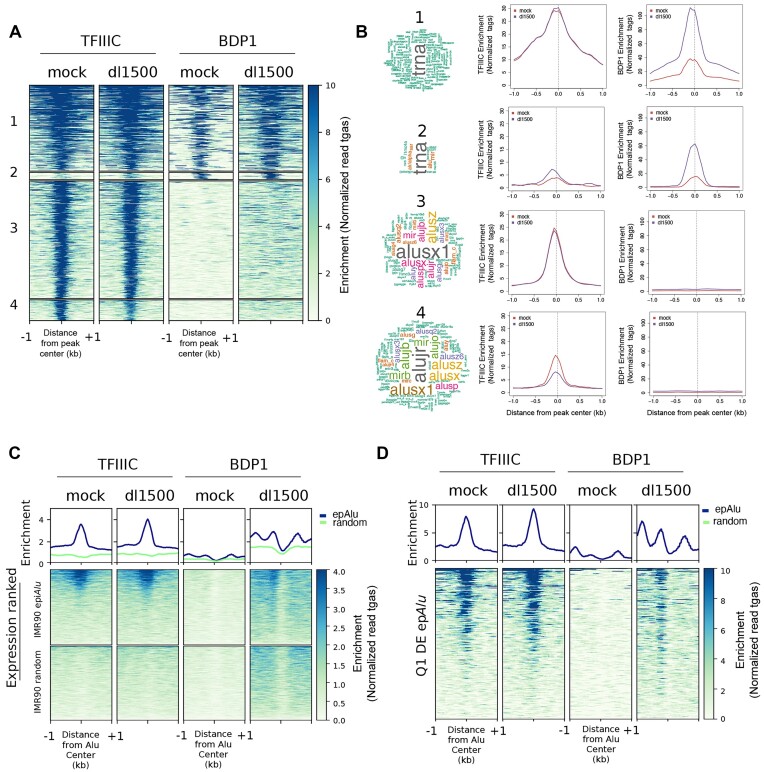
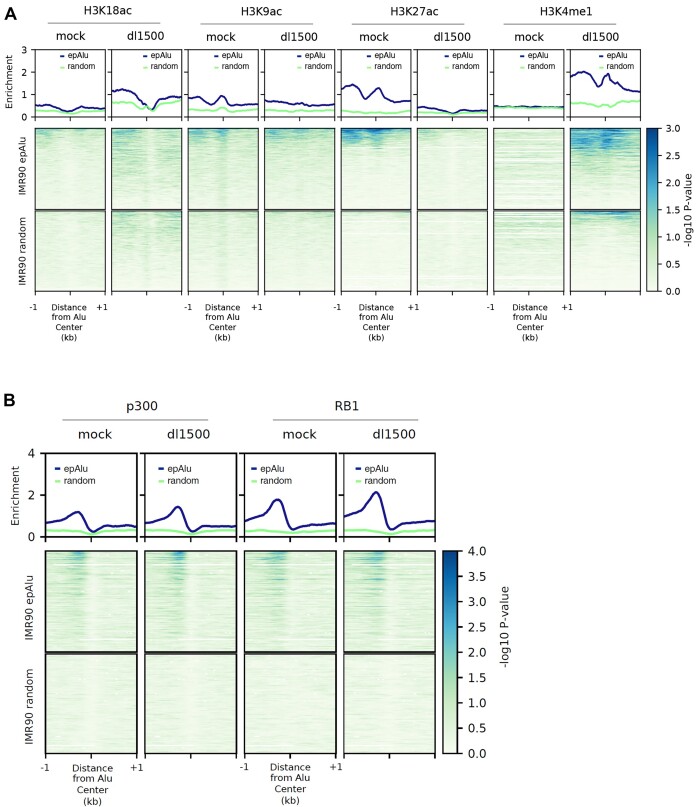
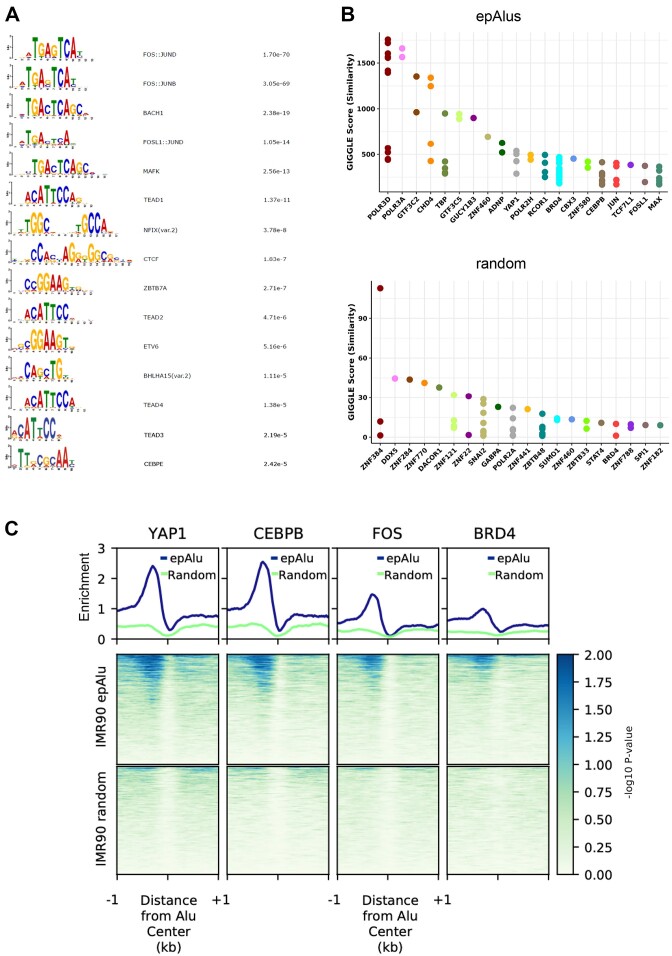
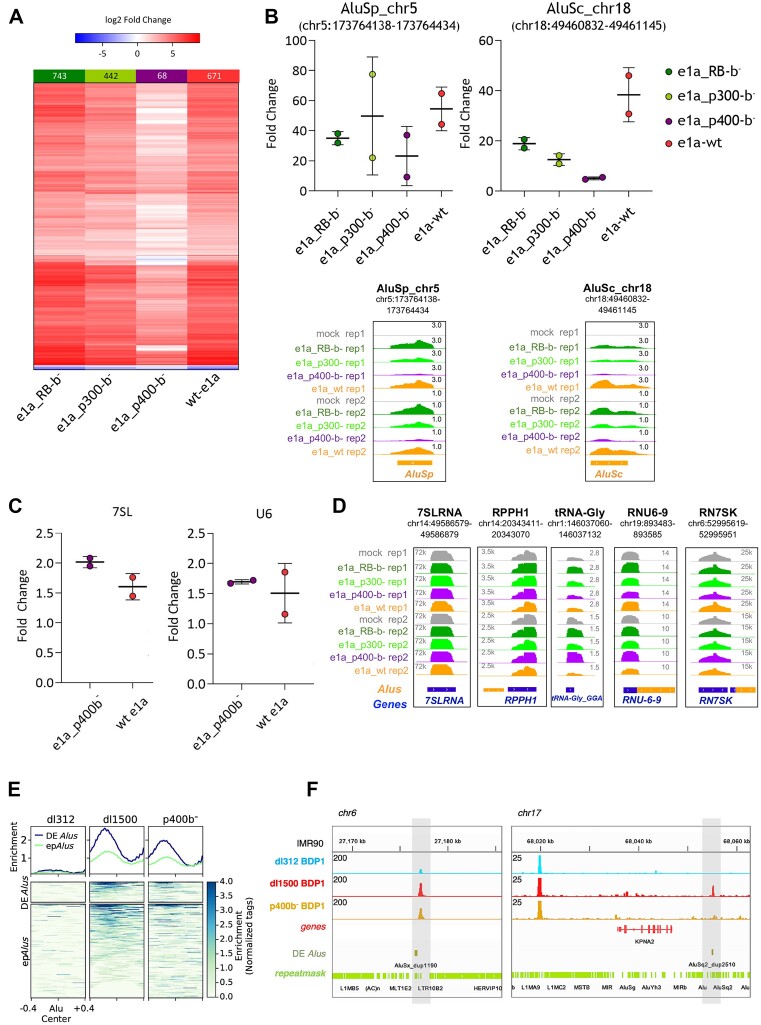
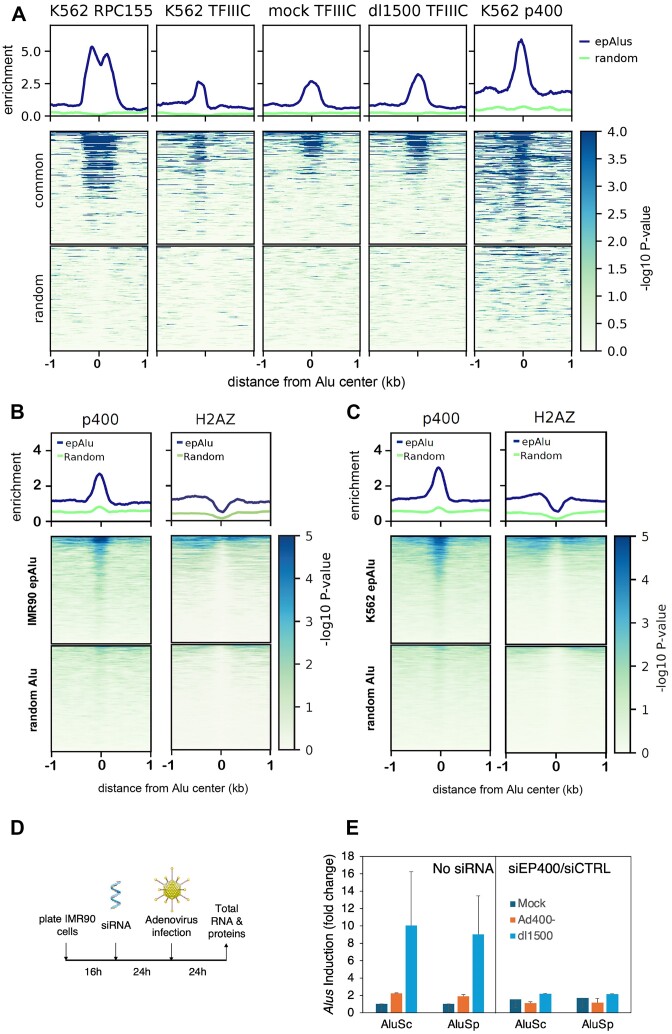
Alu retrotransposons, which form the largest family of mobile DNA elements in the human genome, have recently come to attention as a potential source of regulatory novelties, most notably by participating in enhancer function. Even though Alu transcription by RNA polymerase III is subjected to tight epigenetic silencing, their expression has long been known to increase in response to various types of stress, including viral infection. Here we show that, in primary human fibroblasts, adenovirus small e1a triggered derepression of hundreds of individual Alus by promoting TFIIIB recruitment by Alu-bound TFIIIC. Epigenome profiling revealed an e1a-induced decrease of H3K27 acetylation and increase of H3K4 monomethylation at derepressed Alus, making them resemble poised enhancers. The enhancer nature of e1a-targeted Alus was confirmed by the enrichment, in their upstream regions, of the EP300/CBP acetyltransferase, EP400 chromatin remodeler and YAP1 and FOS transcription factors. The physical interaction of e1a with EP400 was critical for Alu derepression, which was abrogated upon EP400 ablation. Our data suggest that e1a targets a subset of enhancer Alus whose transcriptional activation, which requires EP400 and is mediated by the e1a-EP400 interaction, may participate in the manipulation of enhancer activity by adenoviruses.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Dieci G., Conti A., Pagano A., Carnevali D.. Identification of RNA polymerase III-transcribed genes in eukaryotic genomes. Biochim. Biophys. Acta. 2013; 1829:296–305. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
