

Granulosa cell metabolism at ovulation correlates with oocyte competence and is disrupted by obesity and aging
- PMID: 39013118
- PMCID: PMC11373349
- DOI: 10.1093/humrep/deae154
Granulosa cell metabolism at ovulation correlates with oocyte competence and is disrupted by obesity and aging
Abstract
Study question: Is oocyte developmental competence associated with changes in granulosa cell (GC) metabolism?
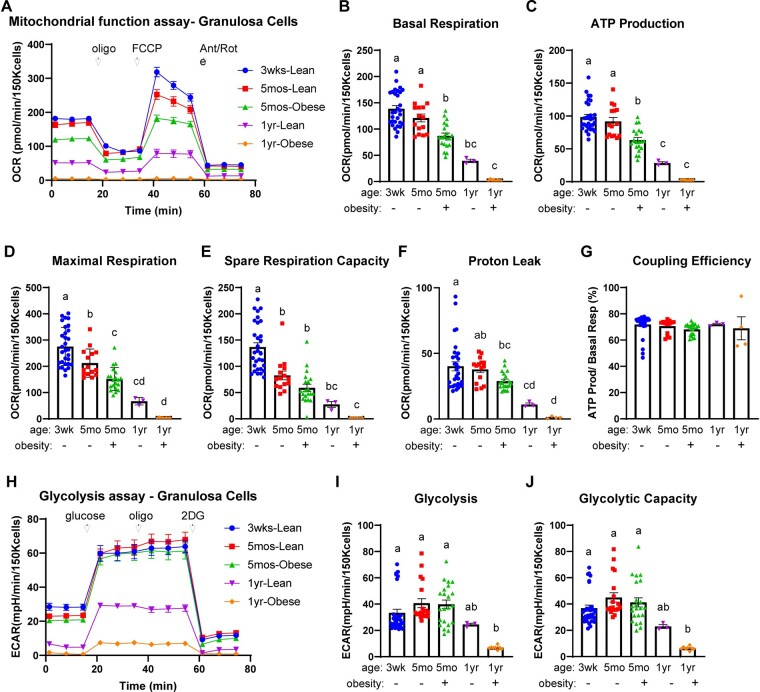
Summary answer: GC metabolism is regulated by the LH surge, altered by obesity and reproductive aging, and, in women, specific metabolic profiles are associated with failed fertilization versus increased blastocyst development.
What is known already: The cellular environment in which an oocyte matures is critical to its future developmental competence. Metabolism is emerging as a potentially important factor; however, relative energy production profiles between GCs and cumulus cells and their use of differential substrates under normal in vivo ovulatory conditions are not well understood.
Study design, size, duration: This study identified metabolic and substrate utilization profiles within ovarian cells in response to the LH surge, using mouse models and GCs of women undergoing gonadotropin-induced oocyte aspiration followed by IVF/ICSI.
Participants/materials, setting, methods: To comprehensively assess follicular energy metabolism, we used real-time metabolic analysis (Seahorse XFe96) to map energy metabolism dynamics (mitochondrial respiration, glycolysis, and fatty acid oxidation) in mouse GCs and cumulus-oocyte complexes (COCs) across a detailed time course in the lead up to ovulation. In parallel, the metabolic profile of GCs was measured in a cohort of 85 women undergoing IVF/ICSI (n = 21 with normal ovarian function; n = 64 with ovarian infertility) and correlated with clinical parameters and cycle outcomes.
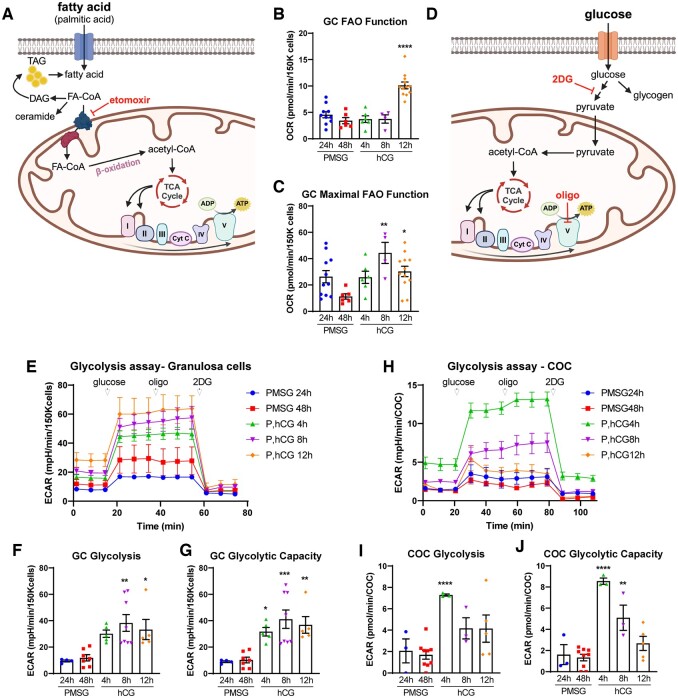
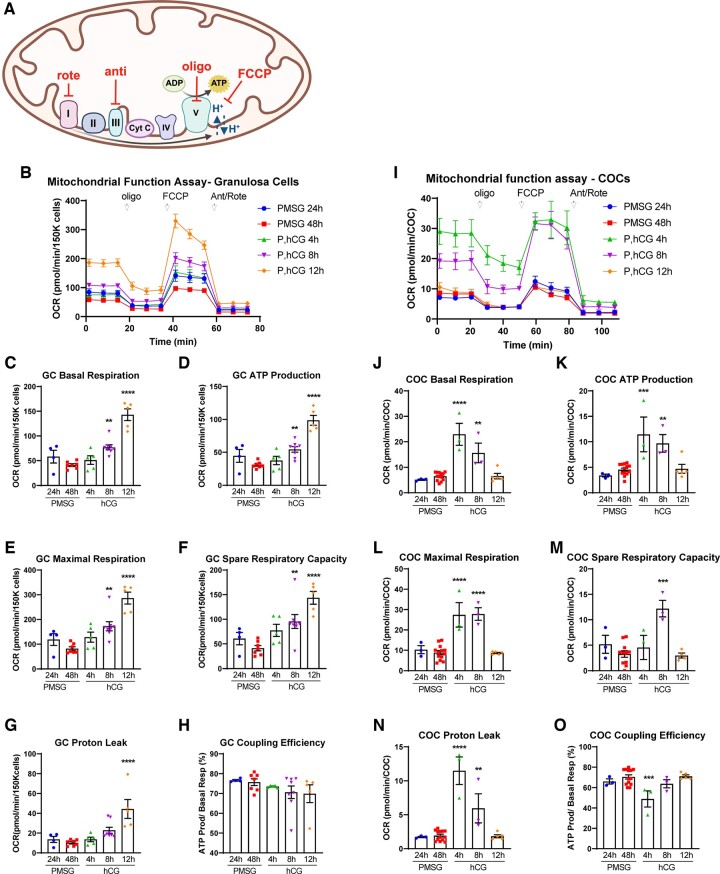
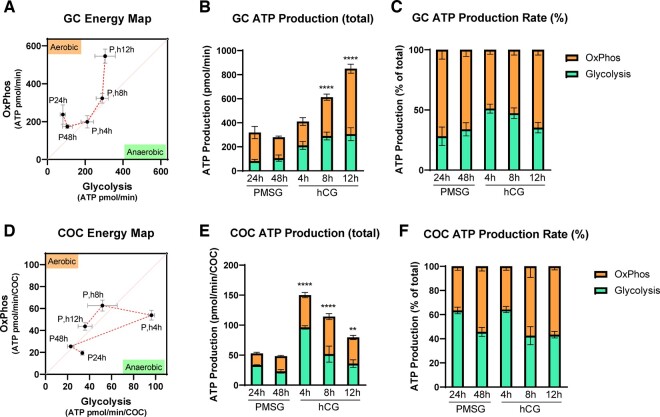
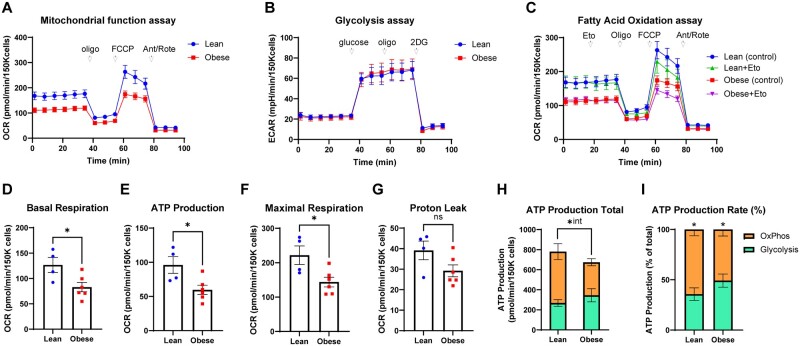
Main results and the role of chance: Our study reveals dynamic changes in GC energy metabolism in response to ovulatory LH, with mitochondrial respiration and glycolysis differentially affected by obesity versus aging, in both mice and women. High respiration in GCs is associated with failed fertilization (P < 0.05) in a subset of women, while glycolytic reserve and mitochondrial ATP production are correlated with on-time development at Day 3 (P < 0.05) and blastocyst formation (P < 0.01) respectively. These data provide new insights into the cellular mechanisms of infertility, by uncovering significant associations between metabolism within the ovarian follicle and oocyte developmental competence.
Limitations, reasons for caution: A larger prospective study is needed before the metabolic markers that were positively and negatively associated with oocyte quality can be used clinically to predict embryo outcomes.
Wider implications of the findings: This study offers new insights into the importance of GC metabolism for subsequent embryonic development and highlights the potential for therapeutic strategies focused on optimizing mitochondrial metabolism to support embryonic development.
Study funding/competing interest(s): National Health and Medical Research Council (Australia). The authors have no competing interests.
Trial registration number: N/A.
Keywords: IVF/ICSI outcome; aging; cumulus–oocyte complex; glycolysis; granulosa cells; metabolism; mitochondria; obesity; ovary; ovulation.
© The Author(s) 2024. Published by Oxford University Press on behalf of European Society of Human Reproduction and Embryology.
Conflict of interest statement
The authors have no conflicts of interest of relevance to this study.
Figures
References
-
- Aardema H, Vos PL, Lolicato F, Roelen BA, Knijn HM, Vaandrager AB, Helms JB, Gadella BM. Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol Reprod 2011;85:62–69. - PubMed
-
- Akison LK, Alvino ER, Dunning KR, Robker RL, Russell DL. Transient invasive migration in mouse cumulus oocyte complexes induced at ovulation by luteinizing hormone. Biol Reprod 2012;86:125. - PubMed
-
- Andreas E, Winstanley YE, Robker RL. Effect of obesity on the ovarian follicular environment and developmental competence of the oocyte. Curr Opin Endocr Metab Res 2021;18:152–158.
-
- Arsov T, Silva DG, O’Bryan MK, Sainsbury A, Lee NJ, Kennedy C, Manji SS, Nelms K, Liu C, Vinuesa CG et al. Fat aussie—a new Alström syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Mol Endocrinol 2006;20:1610–1622. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
