

Myeloid-derived suppressor cells-induced exhaustion of CD8 + T-cell participates in rejection after liver transplantation
- PMID: 39013845
- PMCID: PMC11252260
- DOI: 10.1038/s41419-024-06834-z
Myeloid-derived suppressor cells-induced exhaustion of CD8 + T-cell participates in rejection after liver transplantation
Abstract
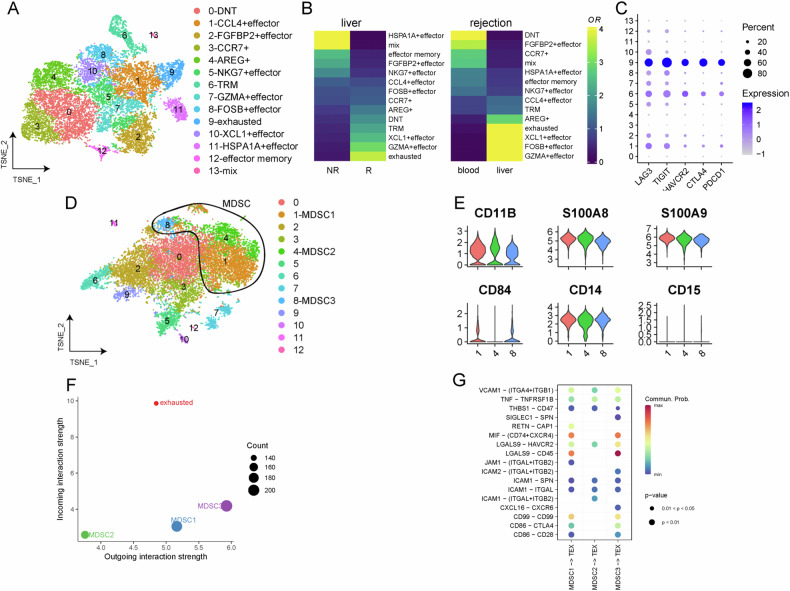
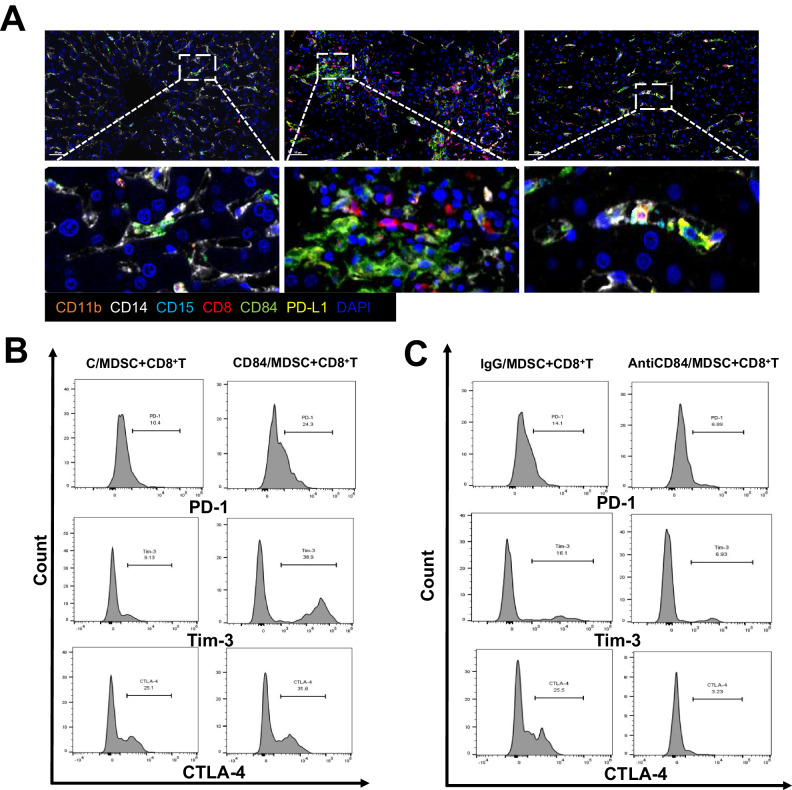
Liver transplantation (LT) rejection remains the most pervasive problem associated with this procedure, while the mechanism involved is still complicated and undefined. One promising solution may involve the use of myeloid-derived suppressor cells (MDSC). However, the immunological mechanisms underlying the effects of MDSC after LT remain unclear. This study is meant to clarify the role MDSCs play after liver transplantation. In this study, we collected liver tissue and peripheral blood mononuclear cells (PBMC) from LT patients showing varying degrees of rejection, as well as liver and spleen tissue samples from mice LT models. These samples were then analyzed using flow cytometry, immunohistochemistry and multiple immunofluorescence. M-MDSCs and CD8 + T-cells extracted from C57/BL6 mice were enriched and cocultured for in vitro experiments. Results, as obtained in both LT patients and LT mice model, revealed that the proportion and frequency of M-MDSC and PD-1 + T-cells increased significantly under conditions associated with a high degree of LT rejection. Within the LT rejection group, our immunofluorescence results showed that a close spatial contiguity was present between PD-1 + T-cells and M-MDSCs in these liver tissue samples and the proportion of CD84/PD-L1 double-positive M-MDSC was greater than that of G-MDSC. There was a positive correlation between the activity of CD84 and immunosuppressive function of M-MDSCs including PD-L1 expression and reactive oxygen species (ROS) production, as demonstrated in our in vitro model. M-MDSCs treated with CD84 protein were able to induce co-cultured CD8 + T-cells to express high levels of exhaustion markers. We found that CD84 regulated M-MDSC function via expression of PD-L1 through activation of the Akt/Stat3 pathway. These results suggest that the capacity for CD84 to regulate M-MDSC induction of CD8 + T-cell exhaustion may play a key role in LT rejection. Such findings provide important, new insights into the mechanisms of tolerance induction in LT.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Lee-Riddle GS, Rocca JP. Low-dose aspirin, rejection, and thrombosis: is it the wonder drug® of liver transplantation? Liver Transpl Publ Am Assoc Study Liver Dis Int Liver Transpl Soc. 2022;28:1825–6. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
