

Selenoprotein W modulates tau homeostasis in an Alzheimer's disease mouse model
- PMID: 39020075
- PMCID: PMC11255228
- DOI: 10.1038/s42003-024-06572-0
Selenoprotein W modulates tau homeostasis in an Alzheimer's disease mouse model
Abstract
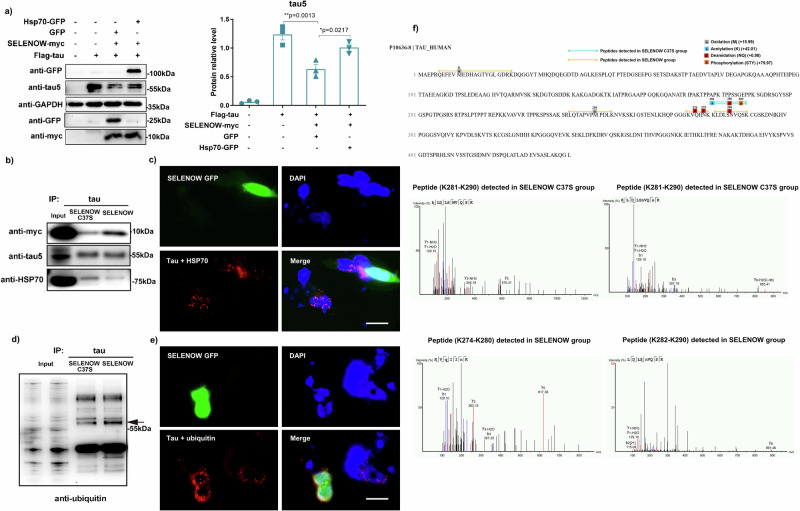
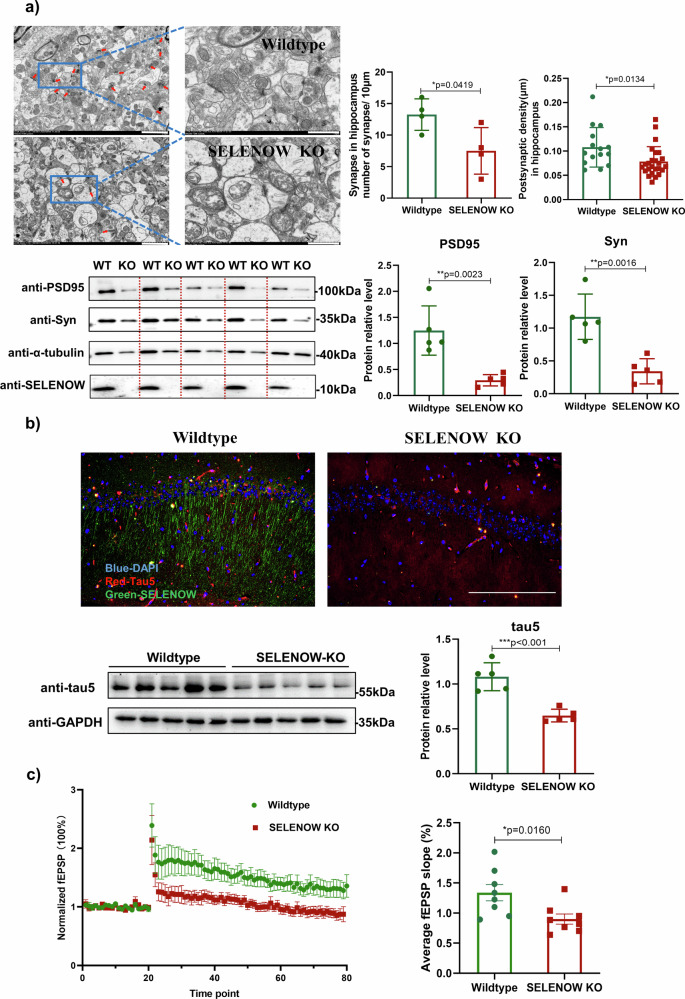
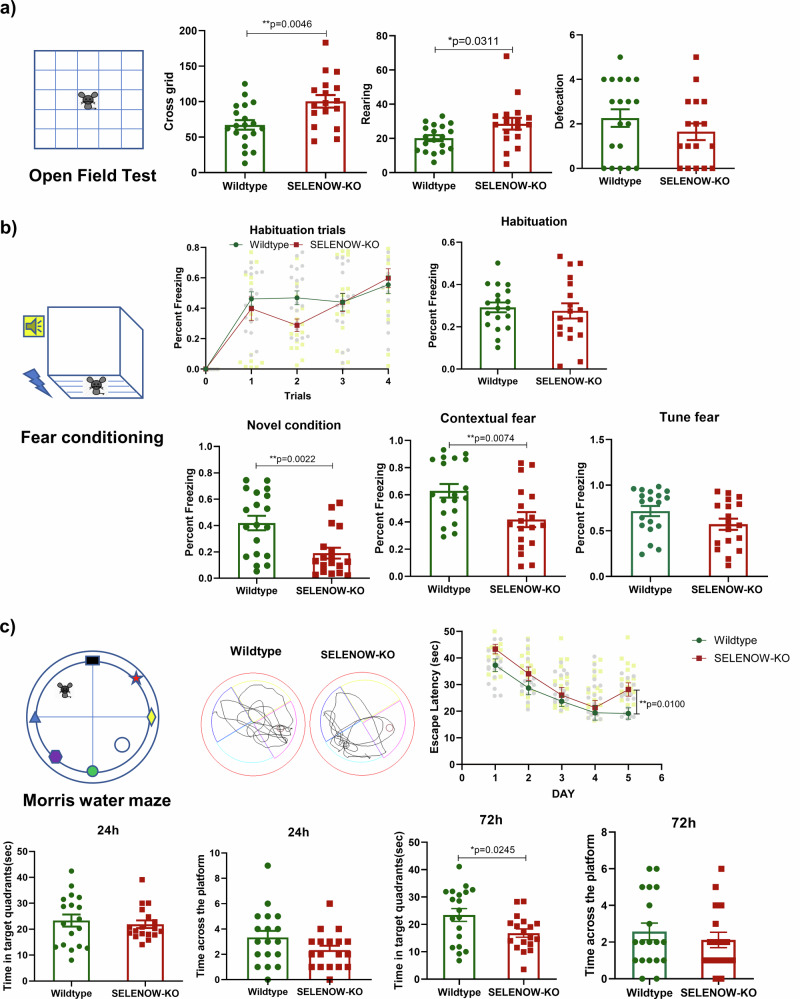
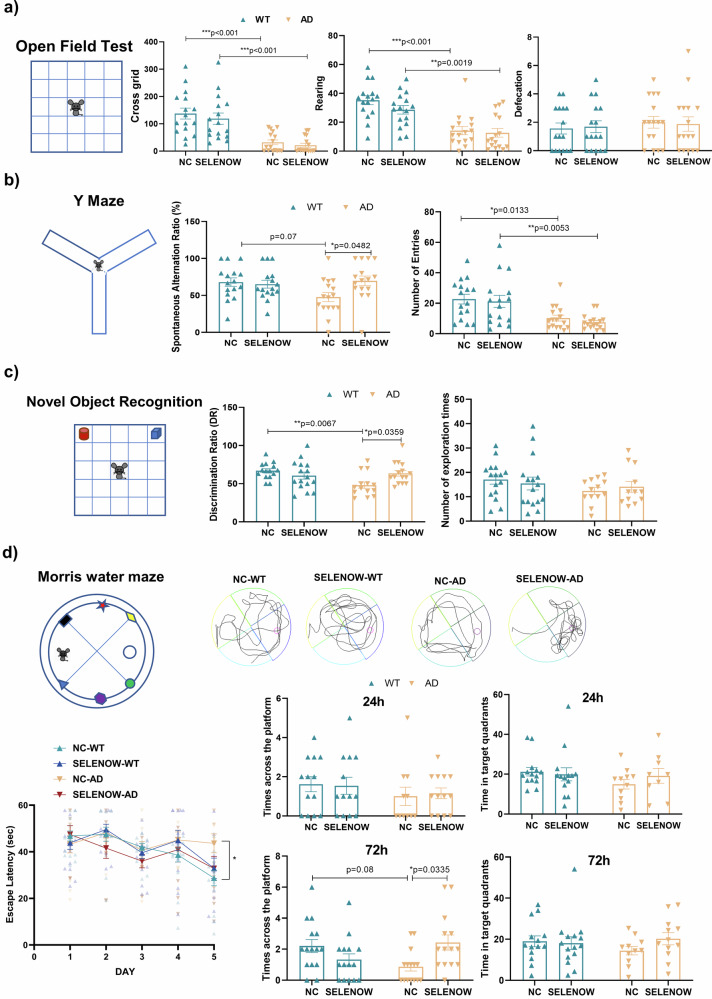
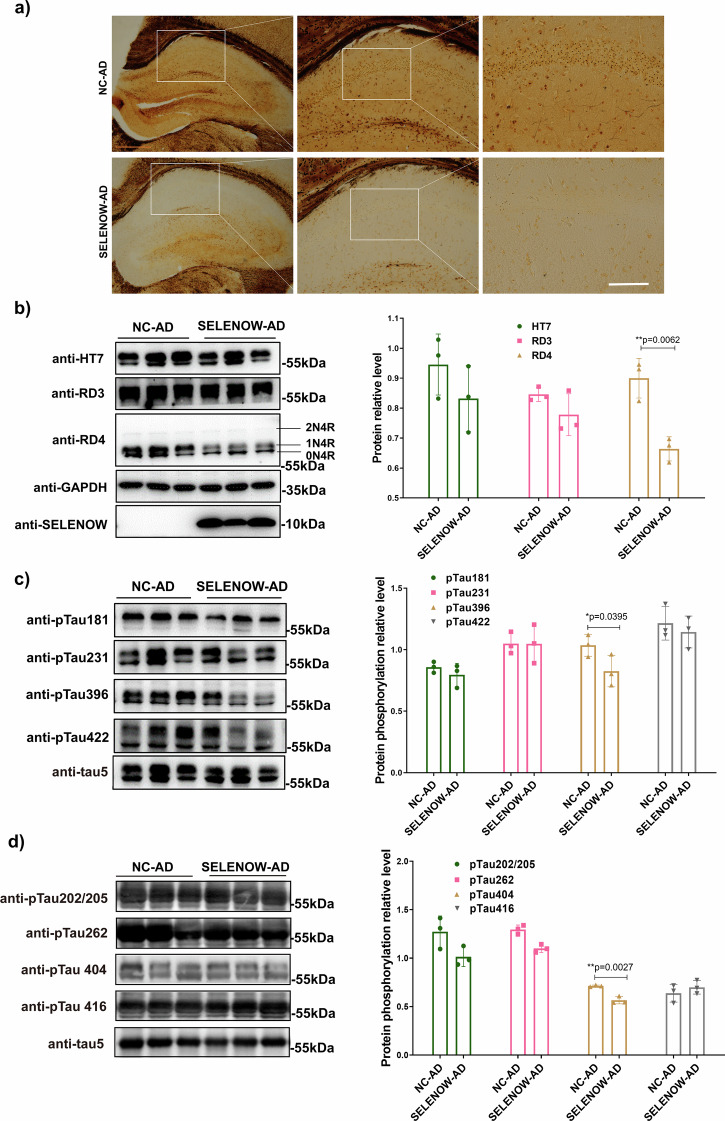
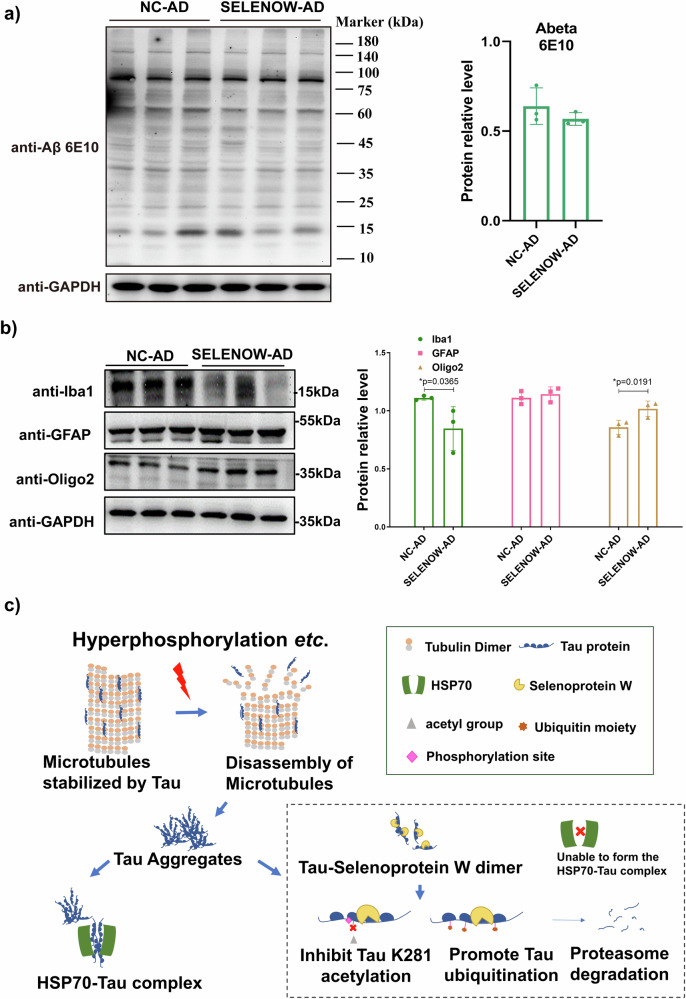
Lower selenium levels are observed in Alzheimer's disease (AD) brains, while supplementation shows multiple benefits. Selenoprotein W (SELENOW) is sensitive to selenium changes and binds to tau, reducing tau accumulation. However, whether restoration of SELENOW has any protective effect in AD models and its underlying mechanism remain unknown. Here, we confirm the association between SELENOW downregulation and tau pathology, revealing SELENOW's role in promoting tau degradation through the ubiquitin‒proteasome system. SELENOW competes with Hsp70 to interact with tau, promoting its ubiquitination and inhibiting tau acetylation at K281. SELENOW deficiency leads to synaptic defects, tau dysregulation and impaired long-term potentiation, resulting in memory deficits in mice. Conversely, SELENOW overexpression in the triple transgenic AD mice ameliorates memory impairment and tau-related pathologies, featuring decreased 4-repeat tau isoform, phosphorylation at Ser396 and Ser404, neurofibrillary tangles and neuroinflammation. Thus, SELENOW contributes to the regulation of tau homeostasis and synaptic maintenance, implicating its potential role in AD.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
