

Acute, chronic and conditioned effects of intranasal oxytocin in the mu-opioid receptor knockout mouse model of autism: Social context matters
- PMID: 39020142
- PMCID: PMC11473707
- DOI: 10.1038/s41386-024-01915-1
Acute, chronic and conditioned effects of intranasal oxytocin in the mu-opioid receptor knockout mouse model of autism: Social context matters
Abstract
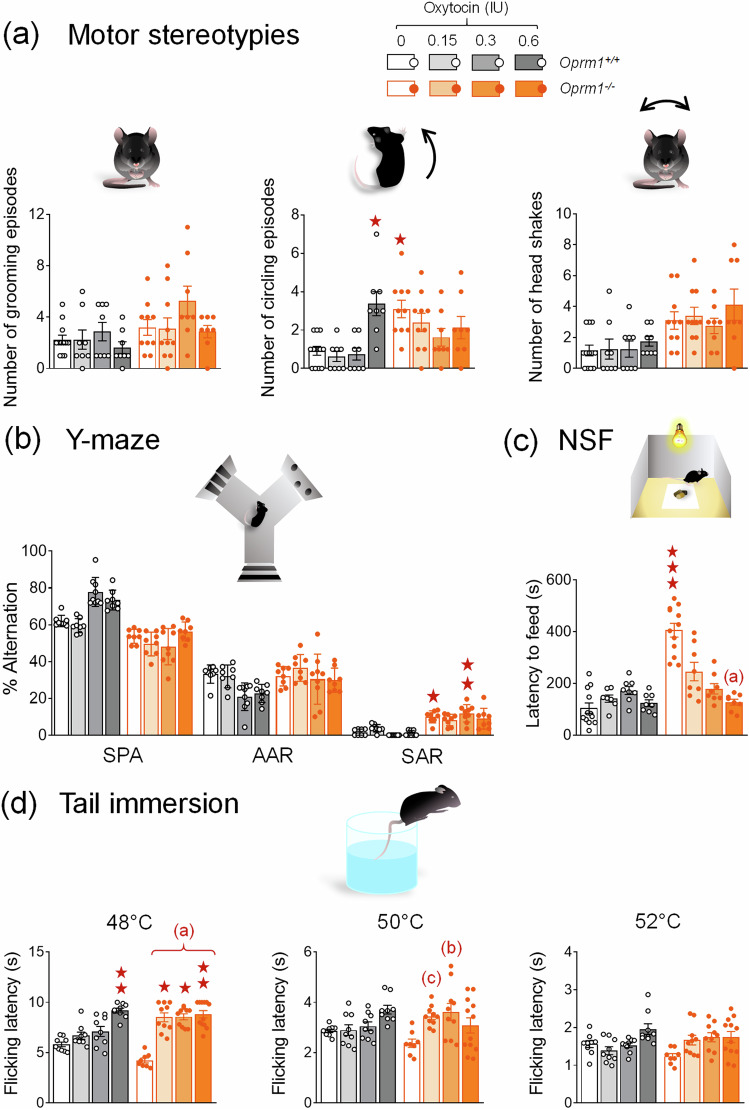
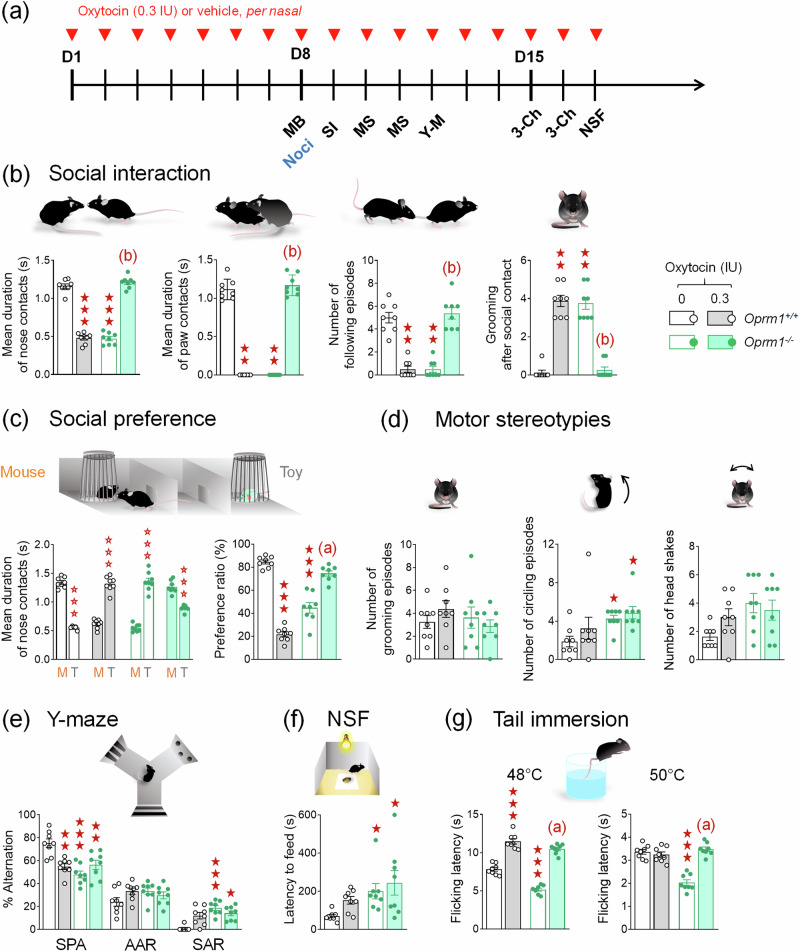
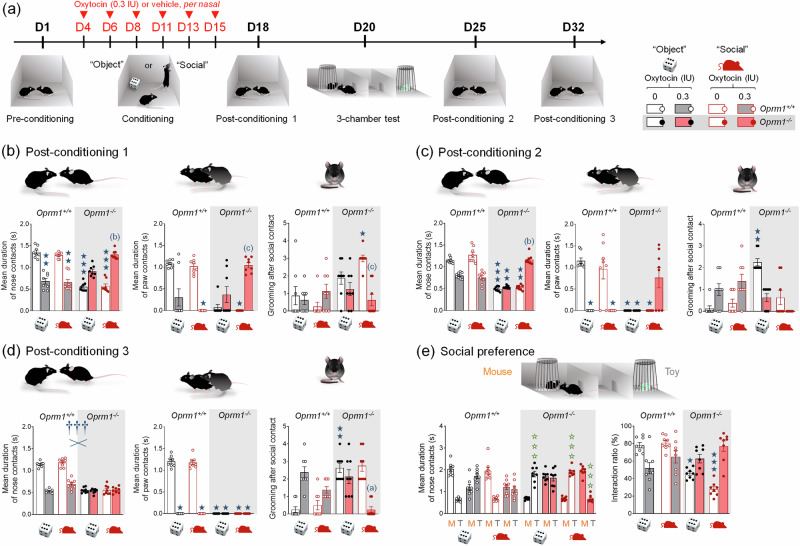
Autism Spectrum Disorders (ASD) are neurodevelopmental disorders whose diagnosis relies on deficient social interaction and communication together with repetitive behaviours. Multiple studies have highlighted the potential of oxytocin (OT) to ameliorate behavioural abnormalities in animal models and subjects with ASD. Clinical trials, however, yielded disappointing results. Our study aimed at assessing the behavioural effects of different regimens of OT administration in the Oprm1 null mouse model of ASD. We assessed the effects of intranasal OT injected once at different doses (0.15, 0.3, and 0.6 IU) and time points (5, 15, and 30 min) following administration, or chronically, on ASD-related behaviours (social interaction and preference, stereotypies, anxiety, nociception) in Oprm1+/+ and Oprm1-/- mice. We then tested whether pairing intranasal OT injection with social experience would influence its outcome on ASD-like symptoms, and measured gene expression in the reward/social circuit. Acute intranasal OT at 0.3 IU improved social behaviour in Oprm1-/- mice 5 min after administration, with limited effects on non-social behaviours. Chronic (8-17 days) OT maintained rescuing effects in Oprm1 null mice but was deleterious in wild-type mice. Finally, improvements in the social behaviour of Oprm1-/- mice were greater and longer lasting when OT was administered in a social context. Under these conditions, the expression of OT and vasopressin receptor genes, as well as marker genes of striatal projection neurons, was suppressed. We detected no sex difference in OT effects. Our results highlight the importance of considering dosage and social context when evaluating the effects of OT treatment in ASD.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- APA. Diagnostic and statistical manual of mental disorders. 2013 5th ed. ed.: Washington, DC.
-
- Johnson CP, Myers SM. Identification and evaluation of children with autism spectrum disorders. Pediatrics. 2007;120:1183–215. - PubMed
-
- Lai MC, Lombardo MV, Baron-Cohen S. Autism. Lancet. 2014;383:896–910. - PubMed
-
- Ramaswami G, Geschwind DH. Genetics of autism spectrum disorder. Handb Clin Neurol. 2018;147:321–29. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
