

Renin-angiotensin system-mediated nitric oxide signaling in podocytes
- PMID: 39024356
- PMCID: PMC11460333
- DOI: 10.1152/ajprenal.00316.2023
Renin-angiotensin system-mediated nitric oxide signaling in podocytes
Abstract
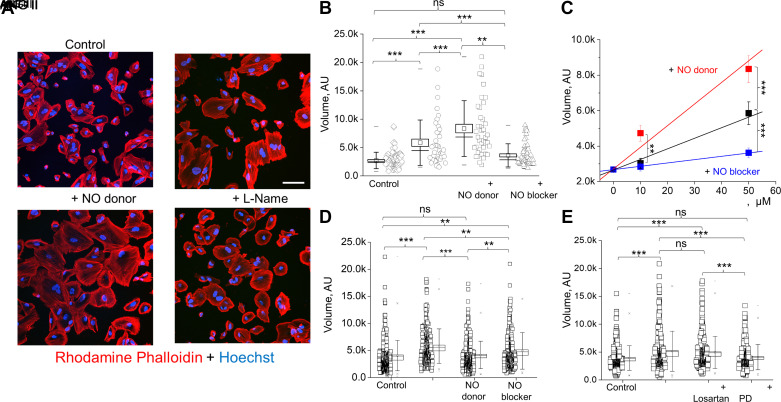
Nitric oxide (NO) is widely recognized for its role in regulating renal function and blood pressure. However, the precise mechanisms by which NO affects renal epithelial cells remain understudied. Our previous research has shown that NO signaling in glomerular podocytes can be initiated by Angiotensin II (ANG II) but not by ATP. This study aims to elucidate the crucial interplay between the renin-angiotensin system (RAS) and NO production in podocytes. To conduct our research, we used cultured human podocytes and freshly isolated rat glomeruli. A variety of RAS peptides were used, alongside confocal microscopy, to detect NO production and NO/Ca2+ cross talk. Dynamic changes in the podocyte cytoskeleton, mediated by RAS-NO intracellular signaling, were observed using fluorescent labeling for F-actin and scanning probe microscopy. The experiments demonstrated that ANG II and ANG III generated high levels of NO by activating the angiotensin II type 2 receptor (AT2R). We did not detect functional MAS receptor presence in podocytes, and the moderate NO response to ANG 1-7 was also mediated through AT2R. Furthermore, NO production impacted intracellular Ca2+ signaling and correlated with an increase in podocyte volume and growth. Scanning probe experiments revealed that AT2R activation and the corresponding NO generation are responsible for the protrusion of podocyte lamellipodia. Taken together, our data indicate that AT2R activation enhances NO production in podocytes and subsequently mediates changes in Ca2+ signaling and podocyte volume dynamics. These mechanisms may play a significant role in both physiological and pathophysiological interactions between the RAS and podocytes.NEW & NOTEWORTHY The renin-angiotensin system plays a crucial role in the production of intracellular nitric oxide within podocytes. This mechanism operates through the activation of the angiotensin II type 2 receptor, leading to dynamic modifications in intracellular calcium levels and the actin filament network. This intricate process is vital for linking the activity of angiotensin receptors to podocyte function.
Keywords: NO bioavailability; angiotensin; cytoskeleton; redox signaling.
Conflict of interest statement
Christine A. Klemens and Alexander Staruschenko are editors of the
Figures








References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous