

This is a preprint.
RBM10 loss induces aberrant splicing of cytoskeletal and extracellular matrix mRNAs and promotes metastatic fitness
- PMID: 39026820
- PMCID: PMC11257529
- DOI: 10.1101/2024.07.09.602730
RBM10 loss induces aberrant splicing of cytoskeletal and extracellular matrix mRNAs and promotes metastatic fitness
Update in
-
RBM10 loss promotes metastases by aberrant splicing of cytoskeletal and extracellular matrix mRNAs.J Exp Med. 2025 May 5;222(5):e20241029. doi: 10.1084/jem.20241029. Epub 2025 Feb 24. J Exp Med. 2025. PMID: 39992626
Abstract
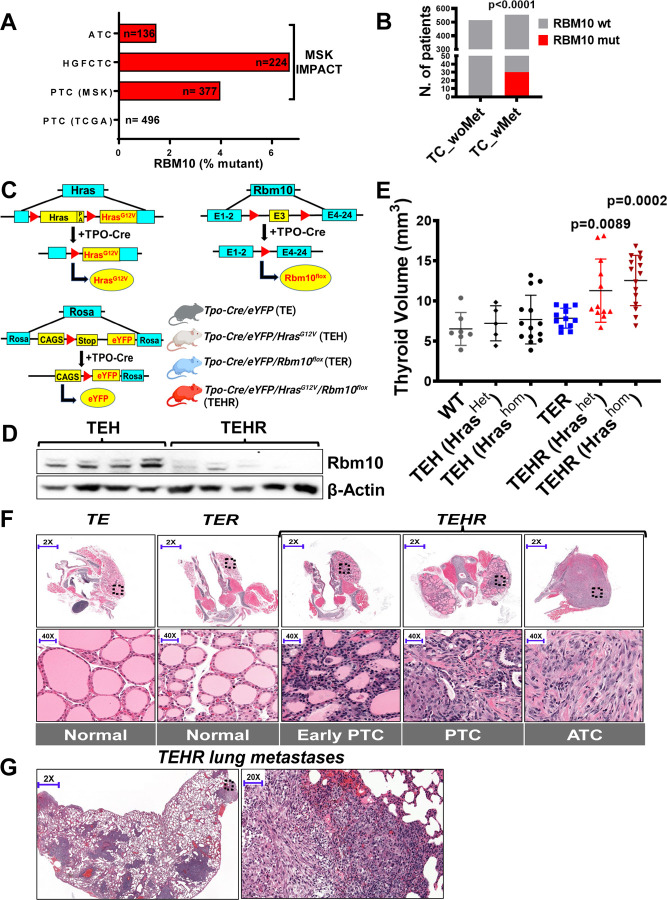
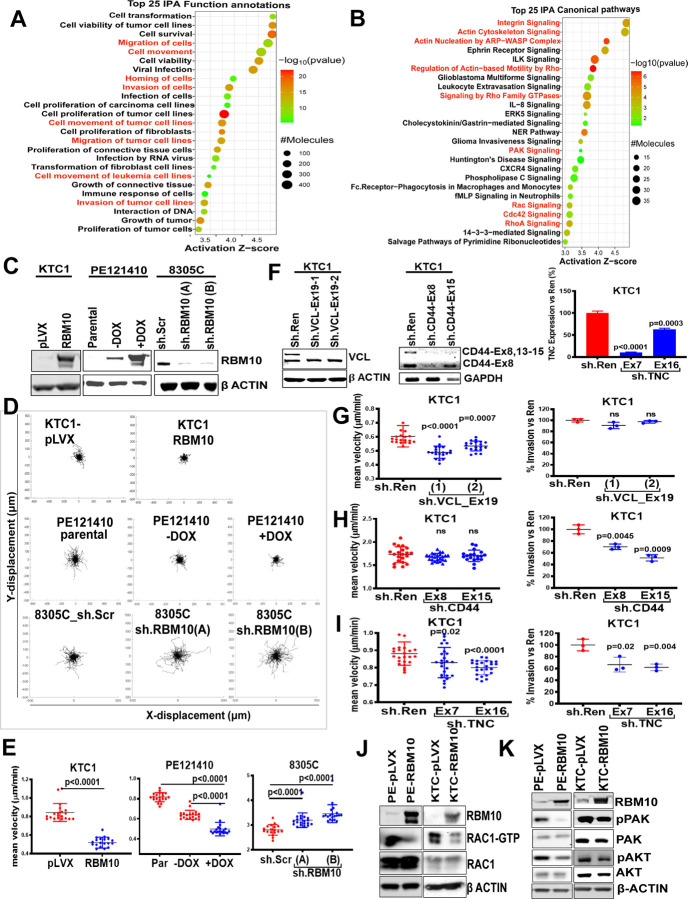
RBM10 modulates transcriptome-wide cassette exon splicing. Loss-of-function RBM10 mutations are enriched in thyroid cancers with distant metastases. Analysis of transcriptomes and genes mis-spliced by RBM10 loss showed pro-migratory and RHO/RAC signaling signatures. RBM10 loss increases cell velocity. Cytoskeletal and ECM transcripts subject to exon-inclusion events included vinculin (VCL), tenascin C (TNC) and CD44. Knockdown of the VCL exon inclusion transcript in RBM10-null cells reduced cell velocity, whereas knockdown of TNC and CD44 exon-inclusion isoforms reduced invasiveness. RAC1-GTP levels were increased in RBM10-null cells. Mouse Hras G12V /Rbm1O KO thyrocytes develop metastases that are reversed by RBM10 or by combined knockdown of VCL, CD44 and TNC inclusion isoforms. Thus, RBM10 loss generates exon inclusions in transcripts regulating ECM-cytoskeletal interactions, leading to RAC1 activation and metastatic competency. Moreover, a CRISPR-Cas9 screen for synthetic lethality with RBM10 loss identified NFkB effectors as central to viability, providing a therapeutic target for these lethal thyroid cancers.
Keywords: CD44; RBM10; TNC; cassette-exon splicing; cell migration; cytoskeletal and ECM remodeling genes; meta-vinculin; metastases; thyroid cancer.
Conflict of interest statement
J.A.F. is a co-inventor of intellectual property focused on HRAS as a biomarker for treating cancer using tipifarnib which has been licensed by MSK to Kura Oncology. J.A.F. received prior research funding from Eisai and was a former consultant for LOXO Oncology, both unrelated to the current manuscript. B.R.U. and J.A.K are co-inventors of intellectual property (HRAS as a biomarker of tipifarnib efficacy) that has been licensed by MSK to Kura Oncology. O.A.-W. has served as a consultant for H3B Biomedicine, Foundation Medicine Inc., Merck, Prelude Therapeutics, and Janssen, and is on the Scientific Advisory Board of Envisagenics Inc., AIChemy, Harmonic Discovery Inc., and Pfizer Boulder. O.A.-W. has received prior research funding from H3B Biomedicine, Nurix Therapeutics, Minovia Therapeutics, and LOXO Oncology unrelated to the current manuscript. R.K.B. is an inventor on patent applications filed by Fred Hutchinson Cancer Center related to modulating splicing for cancer therapy. R.K.B. and O.A.-W. are founders and scientific advisors of Codify Therapeutics, hold equity in this company and receive research support from this company unrelated to the current manuscript. R.K.B. is a founder and scientific advisor of Synthesize Bio and holds equity in this company. A.R.G is currently an Associate Professor of Surgery at the University of Sydney, Australia. M.S. is currently employed by Loxo Oncology.The remaining authors declare no competing interests.
Figures



References
-
- ARUFFO A., STAMENKOVIC I., MELNICK M., UNDERHILL C. B. & SEED B. 1990. CD44 is the principal cell surface receptor for hyaluronate. Cell, 61, 1303–13. - PubMed
-
- BAGHERI-YARMAND R., BUSAIDY N. L., MCBEATH E., DANYSH B. P., EVANS K. W., MOSS T. J., AKCAKANAT A., NG P. K. S., KNIPPLER C. M., GOLDEN J. A., WILLIAMS M. D., MULTANI A. S., CABANILLAS M. E., SHAW K. R., MERIC-BERNSTAM F., SHAH M. H., RINGEL M. D. & HOFMANN M. C. 2021. RAC1 Alterations Induce Acquired Dabrafenib Resistance in Association with Anaplastic Transformation in a Papillary Thyroid Cancer Patient. Cancers (Basel), 13. - PMC - PubMed
-
- BECHARA E. G., SEBESTYEN E., BERNARDIS I., EYRAS E. & VALCARCEL J. 2013. RBM5, 6, and 10 differentially regulate NUMB alternative splicing to control cancer cell proliferation. Mol Cell, 52, 720–33. - PubMed
-
- BELKIN A. M., ORNATSKY O. I., KABAKOV A. E., GLUKHOVA M. A. & KOTELIANSKY V. E. 1988b. Diversity of vinculin/meta-vinculin in human tissues and cultivated cells. Expression of muscle specific variants of vinculin in human aorta smooth muscle cells. J Biol Chem, 263, 6631–5. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous