

Protein semisynthesis reveals plasticity in HECT E3 ubiquitin ligase mechanisms
- PMID: 39030419
- PMCID: PMC12087361
- DOI: 10.1038/s41557-024-01576-z
Protein semisynthesis reveals plasticity in HECT E3 ubiquitin ligase mechanisms
Abstract
Lys ubiquitination is catalysed by E3 ubiquitin ligases and is central to the regulation of protein stability and cell signalling in normal and disease states. There are gaps in our understanding of E3 mechanisms, and here we use protein semisynthesis, chemical rescue, microscale thermophoresis and other biochemical approaches to dissect the role of catalytic base/acid function and conformational interconversion in HECT-domain E3 catalysis. We demonstrate that there is plasticity in the use of the terminal side chain or backbone carboxylate for proton transfer in HECT E3 ubiquitin ligase reactions, with yeast Rsp5 orthologues appearing to be possible evolutionary intermediates. We also show that the HECT-domain ubiquitin covalent intermediate appears to eject the E2 conjugating enzyme, promoting catalytic turnover. These findings provide key mechanistic insights into how protein ubiquitination occurs and provide a framework for understanding E3 functions and regulation.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures







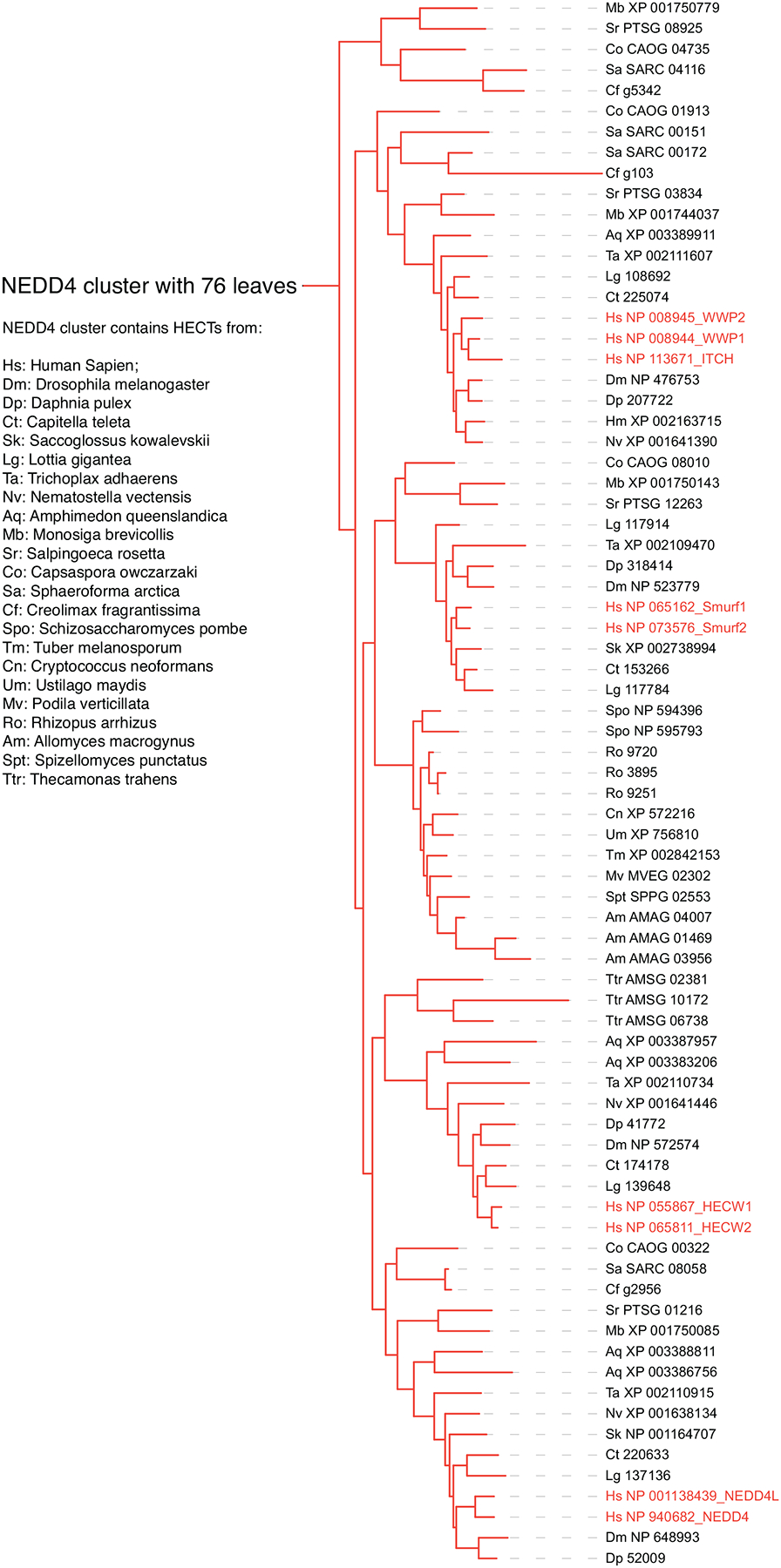
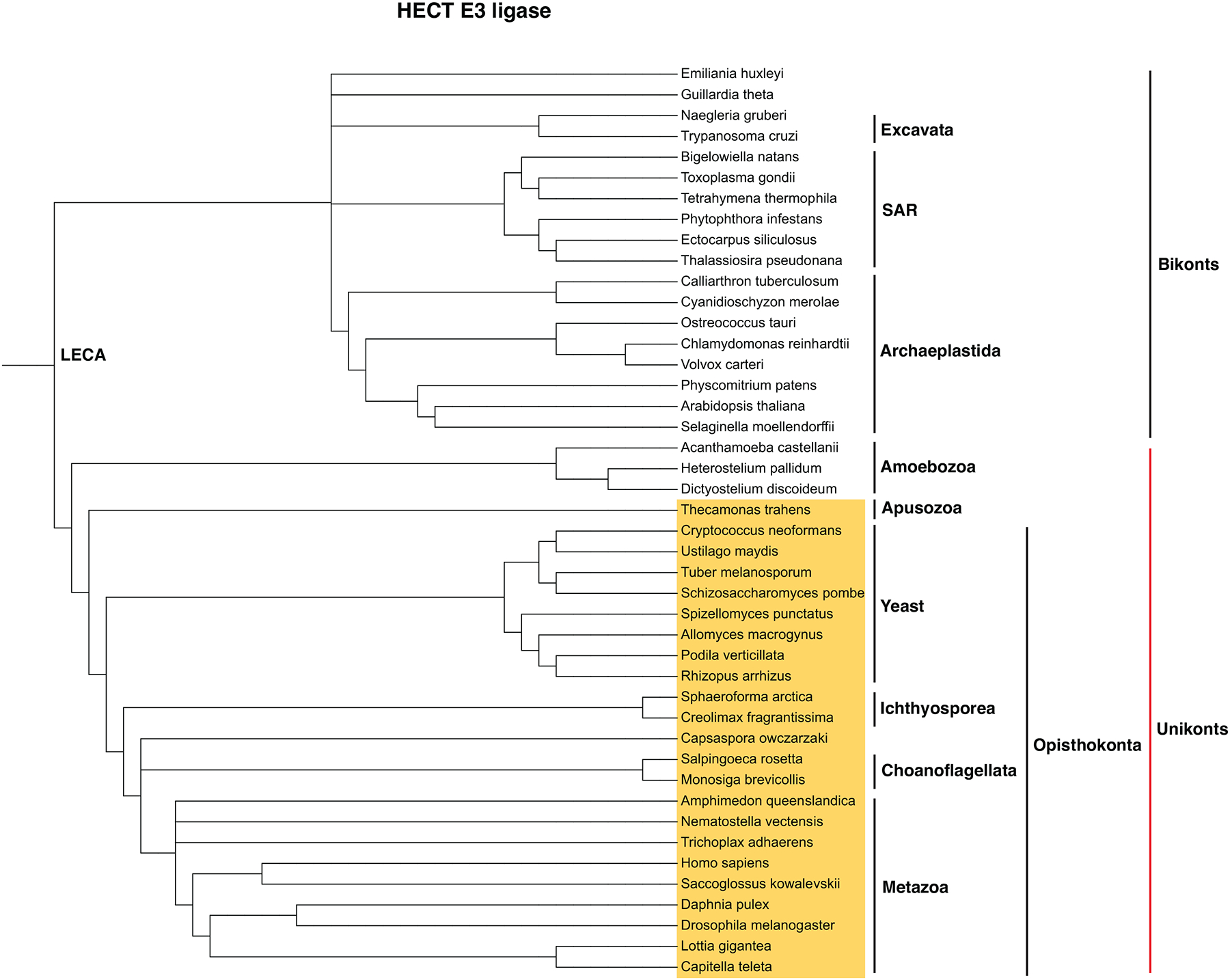





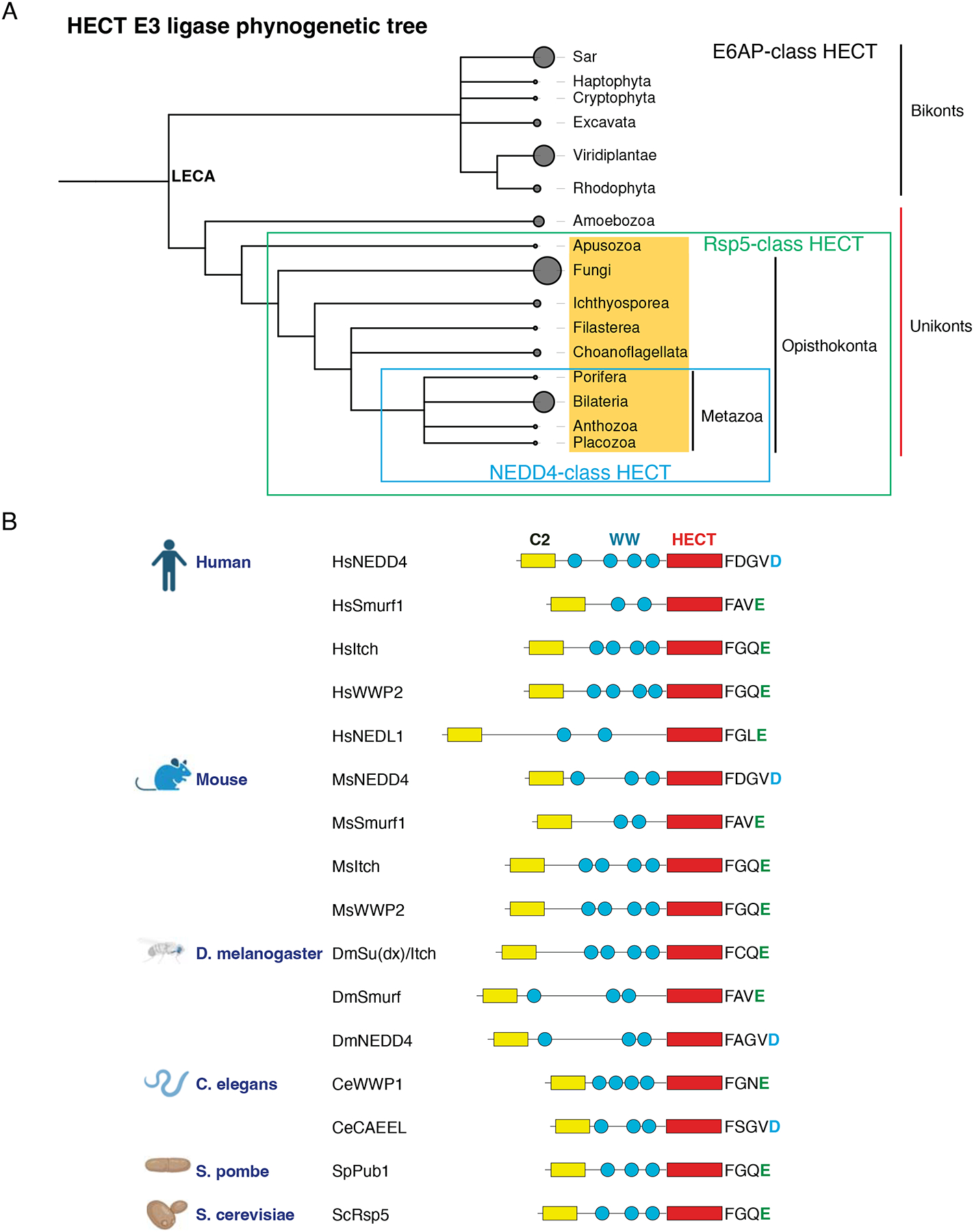
References
-
- Komander D & Rape M The ubiquitin code. Annual review of biochemistry 81, 203–229 (2012). - PubMed
MeSH terms
Substances
Grants and funding
- R01 CA074305/CA/NCI NIH HHS/United States
- CA74305/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- GM R35 149229/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM062437/GM/NIGMS NIH HHS/United States
- 826614/American Heart Association (American Heart Association, Inc.)
LinkOut - more resources
Full Text Sources
