

Preclinical studies of gene replacement therapy for CDKL5 deficiency disorder
- PMID: 39033321
- PMCID: PMC11489525
- DOI: 10.1016/j.ymthe.2024.07.012
Preclinical studies of gene replacement therapy for CDKL5 deficiency disorder
Abstract
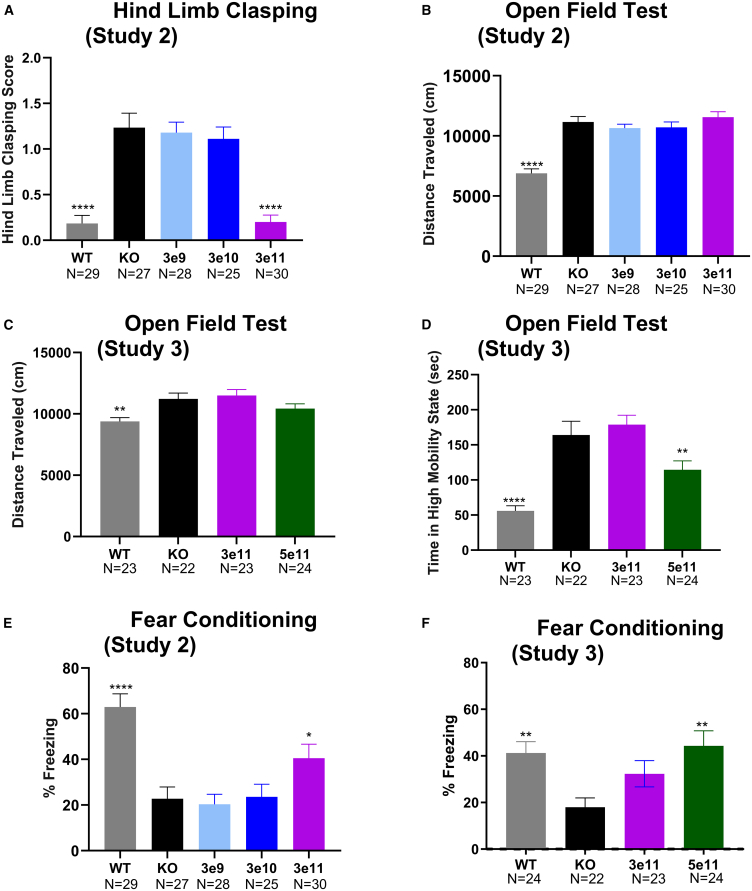
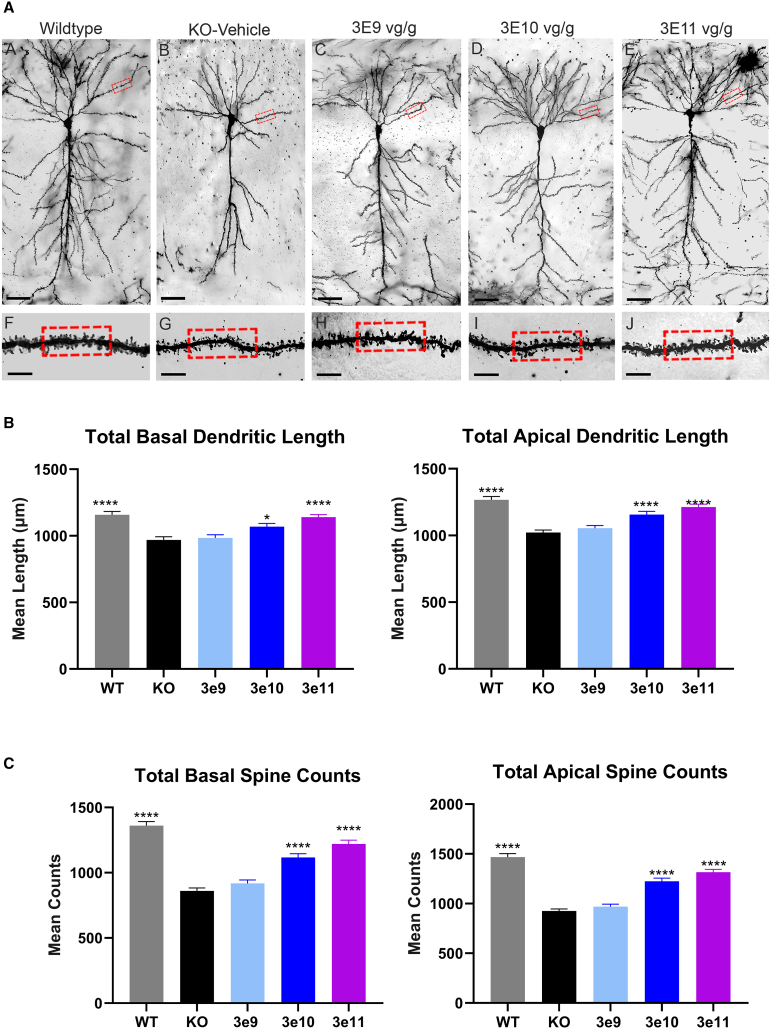
Cyclin-dependent kinase-like 5 (CDKL5) deficiency disorder (CDD) is a rare neurodevelopmental disorder caused by a mutation in the X-linked CDKL5 gene. CDKL5 is a serine/threonine kinase that is critical for axon outgrowth and dendritic morphogenesis as well as synapse formation, maturation, and maintenance. This disorder is characterized by early-onset epilepsy, hypotonia, and failure to reach cognitive and motor developmental milestones. Because the disease is monogenic, delivery of the CDKL5 gene to the brain of patients should provide clinical benefit. To this end, we designed a gene therapy vector, adeno-associated virus (AAV)9.Syn.hCDKL5, in which human CDKL5 gene expression is driven by the synapsin promoter. In biodistribution studies conducted in mice, intracerebroventricular (i.c.v.) injection resulted in broader, more optimal biodistribution than did intra-cisterna magna (i.c.m.) delivery. AAV9.Syn.hCDKL5 treatment increased phosphorylation of EB2, a bona fide CDKL5 substrate, demonstrating biological activity in vivo. Our data provide proof of concept that i.c.v. delivery of AAV9.Syn.hCDKL5 to neonatal male Cdkl5 knockout mice reduces pathology and reduces aberrant behavior. Functional improvements were seen at doses of 3e11 to 5e11 vector genomes/g brain, which resulted in transfection of ≥50% of the neurons. Functional improvements were not seen at lower doses, suggesting a requirement for broad distribution for efficacy.
Keywords: AAV9; CDD; CDKL5; biodistribution; drug discovery and development; gene therapy; molecular therapeutics; mouse models of disease; pharmacodynamics; route of administration.
Copyright © 2024 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.V., J.N., J.G., J.S., P.L., M.P., S.D., L.C., Y.V., M.J.K., L.P., E.F., S.S., B.R., P.Y., C.S., J.P., K.S., S.J., J.L., A.M., E.M.W., Z.W., and M.W. are or were at the time the work employees of PTC Therapeutics. M.C.W. is the President and CEO of NeuroDigiTech and was paid by PTC Therapeutics for this work.
Figures




References
-
- Fehr S., Wong K., Chin R., Williams S., de Klerk N., Forbes D., Krishnaraj R., Christodoulou J., Downs J., Leonard H. Seizure variables and their relationship to genotype and functional abilities in the CDKL5 disorder. Neurology. 2016;87:2206–2213. - PubMed
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
