

Targeted degradation of extracellular mitochondrial aspartyl-tRNA synthetase modulates immune responses
- PMID: 39039092
- PMCID: PMC11263397
- DOI: 10.1038/s41467-024-50031-7
Targeted degradation of extracellular mitochondrial aspartyl-tRNA synthetase modulates immune responses
Abstract
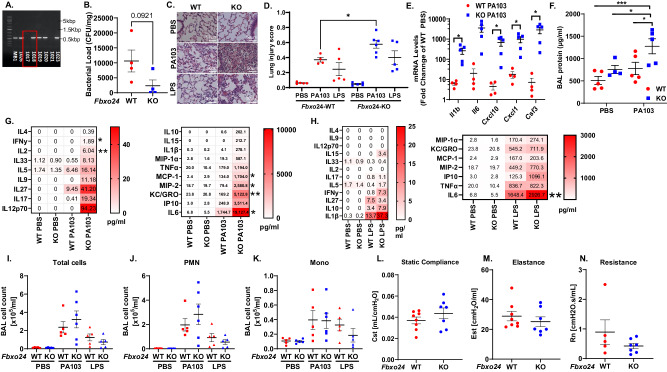
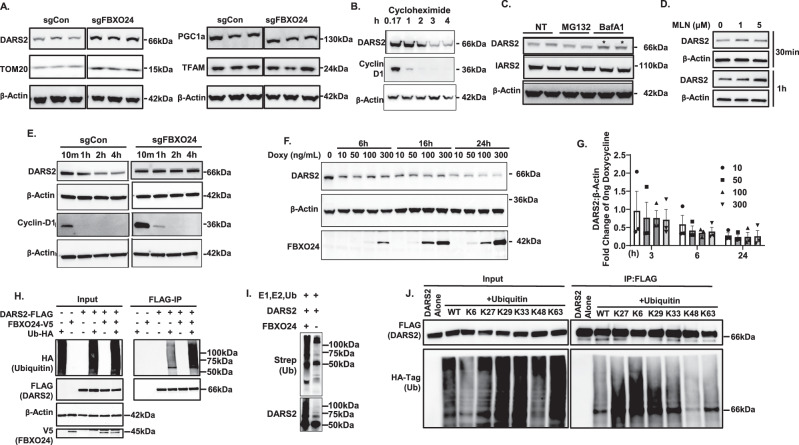
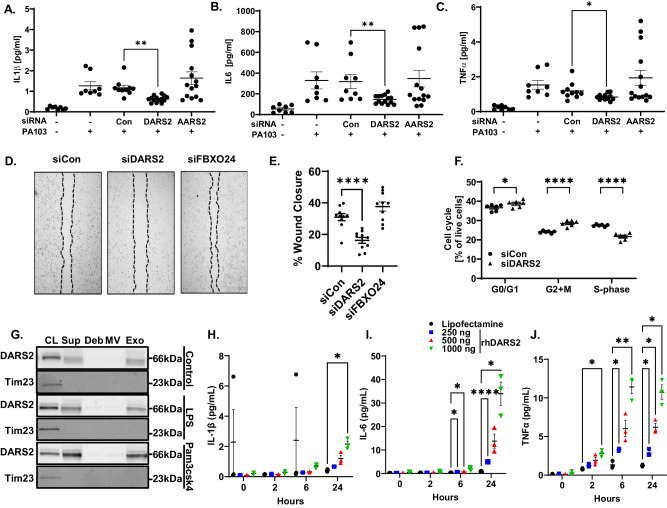
The severity of bacterial pneumonia can be worsened by impaired innate immunity resulting in ineffective pathogen clearance. We describe a mitochondrial protein, aspartyl-tRNA synthetase (DARS2), which is released in circulation during bacterial pneumonia in humans and displays intrinsic innate immune properties and cellular repair properties. DARS2 interacts with a bacterial-induced ubiquitin E3 ligase subunit, FBXO24, which targets the synthetase for ubiquitylation and degradation, a process that is inhibited by DARS2 acetylation. During experimental pneumonia, Fbxo24 knockout mice exhibit elevated DARS2 levels with an increase in pulmonary cellular and cytokine levels. In silico modeling identified an FBXO24 inhibitory compound with immunostimulatory properties which extended DARS2 lifespan in cells. Here, we show a unique biological role for an extracellular, mitochondrially derived enzyme and its molecular control by the ubiquitin apparatus, which may serve as a mechanistic platform to enhance protective host immunity through small molecule discovery.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
- R01HL096376/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 HL097376/HL/NHLBI NIH HHS/United States
- P01HL114453/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K08 HL169725/HL/NHLBI NIH HHS/United States
- R01 HL169586/HL/NHLBI NIH HHS/United States
- R01HL081784/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- P01 HL114453/HL/NHLBI NIH HHS/United States
- P30 CA016058/CA/NCI NIH HHS/United States
- R01 HL096376/HL/NHLBI NIH HHS/United States
- R01 HL081784/HL/NHLBI NIH HHS/United States
- R35 HL139860/HL/NHLBI NIH HHS/United States
- R01HL097376/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
