

Neural-circuit basis of song preference learning in fruit flies
- PMID: 39040064
- PMCID: PMC11260866
- DOI: 10.1016/j.isci.2024.110266
Neural-circuit basis of song preference learning in fruit flies
Abstract
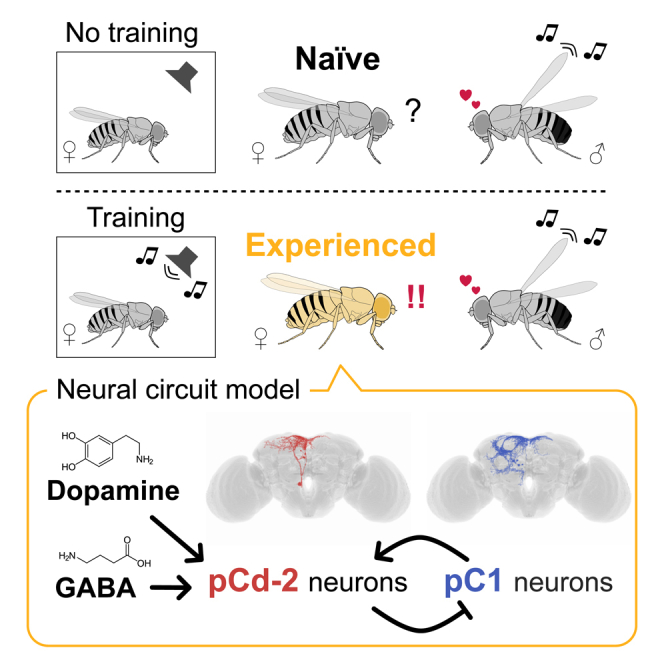
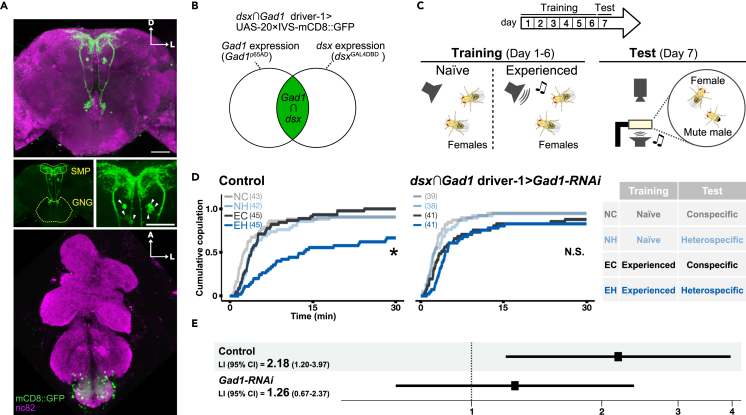
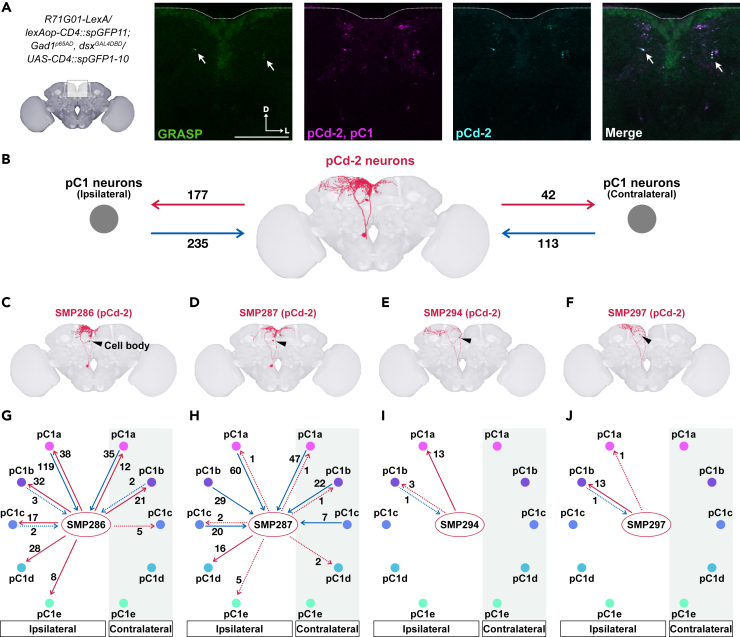
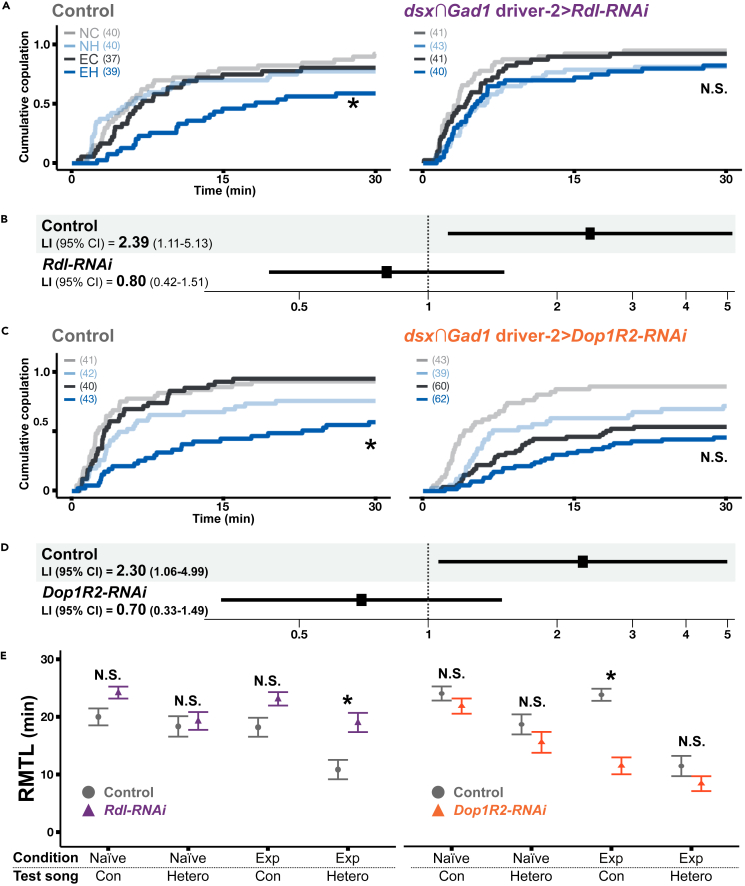
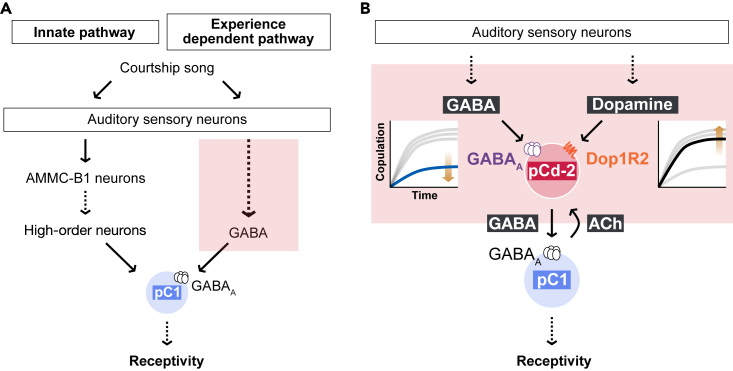
As observed in human language learning and song learning in birds, the fruit fly Drosophila melanogaster changes its auditory behaviors according to prior sound experiences. This phenomenon, known as song preference learning in flies, requires GABAergic input to pC1 neurons in the brain, with these neurons playing a key role in mating behavior. The neural circuit basis of this GABAergic input, however, is not known. Here, we find that GABAergic neurons expressing the sex-determination gene doublesex are necessary for song preference learning. In the brain, only four doublesex-expressing GABAergic neurons exist per hemibrain, identified as pCd-2 neurons. pCd-2 neurons directly, and in many cases mutually, connect with pC1 neurons, suggesting the existence of reciprocal circuits between them. Moreover, GABAergic and dopaminergic inputs to doublesex-expressing GABAergic neurons are necessary for song preference learning. Together, this study provides a neural circuit model that underlies experience-dependent auditory plasticity at a single-cell resolution.
Keywords: Behavioral neuroscience; Cellular neuroscience; Molecular neuroscience.
© 2024 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Gentner T.Q., Margoliash D. In: Acoustic Communication. Simmons A.M., Popper A.N., Fay R.R., editors. Springer-Verlag; 2003. The Neuroethology of Vocal Communication: Perception and Cognition; pp. 324–386. - DOI
-
- Thorpe W.H. The learning of song patterns by birds, with especial reference to the song of the Chaffinch Fringilla coelebs. Ibis. 2008;100:535–570. doi: 10.1111/j.1474-919X.1958.tb07960.x. - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
