

This is a preprint.
A hypothalamic circuit for circadian regulation of corticosterone secretion
- PMID: 39041039
- PMCID: PMC11261983
- DOI: 10.21203/rs.3.rs-4718850/v2
A hypothalamic circuit for circadian regulation of corticosterone secretion
Abstract
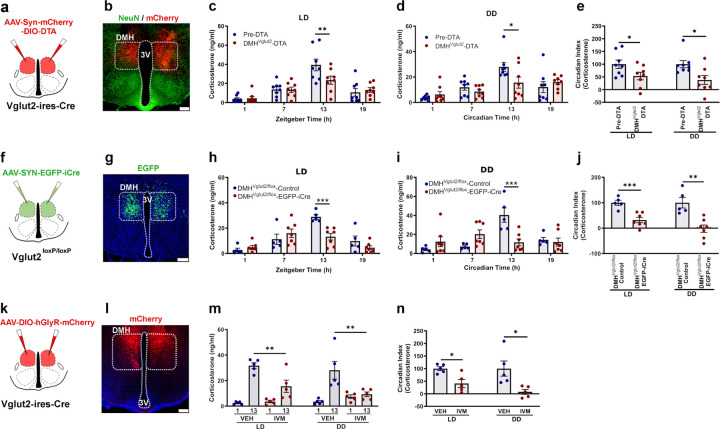
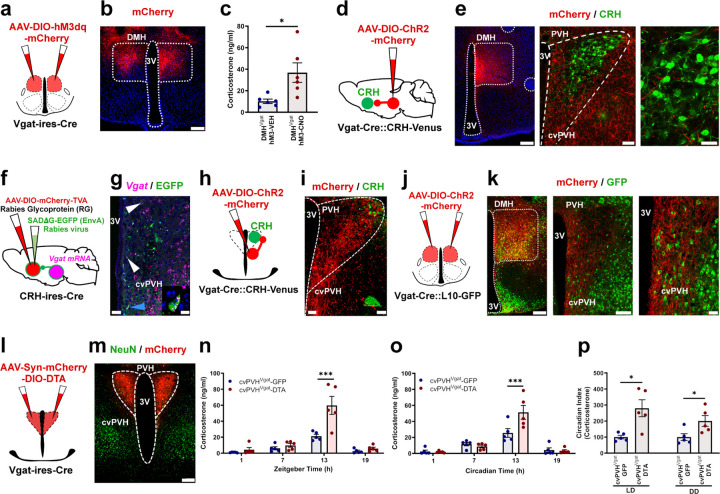
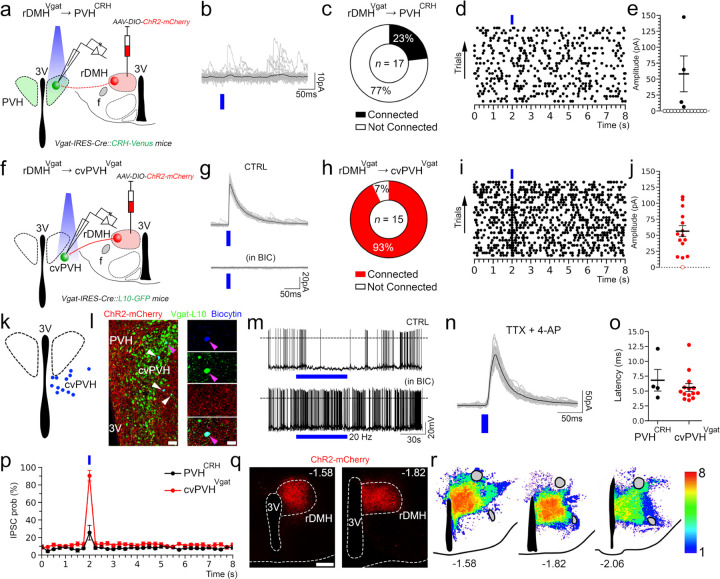
The secretion of cortisol in humans and corticosterone (Cort) in rodents follows a daily rhythm which is important in readying the individual for daily activity. This rhythm is orchestrated by the suprachiasmatic nucleus (SCN), but how it ultimately regulates the circadian rhythm of activity of neurons in the paraventricular nucleus of the hypothalamus that produce corticotropin-releasing hormone (PVHCRH neurons) is not known. We hypothesized that the SCN may exert this influence by projections to the subparaventricular zone (SPZ), which in turn innervates neurons in the dorsomedial nucleus of the hypothalamus (DMH) that regulate PVHCRH neurons. First, we found that ablating SPZVgat neurons eliminates the circadian rhythm of Cort secretion, but that deleting Vgat from them does not, suggesting that they predominantly use some other transmitter. Next, we found that either ablating or acutely inhibiting the DMH glutamatergic (DMHVglut2) neurons resulted in a 40-70% reduction in the daily peak of Cort. Deletion of the Vglut2 gene within the DMH produced a similar effect, highlighting the indispensable role of glutamatergic signaling. Chemogenetic stimulation of DMHVglut2 neurons led to an increase of Cort levels, and optogenetic activation of their terminals in the PVH in hypothalamic slices directly activated PVHCRH neurons through glutamate action on AMPA receptors (the DMHVglut2 → PVHCRH pathway). Similar to the disruption of DMHVglut2 neurons, ablating, inhibiting, or disrupting GABA transmission by DMH GABAergic (DMHVgat) neurons diminished the circadian peak of Cort, particularly under constant darkness conditions. Chemogenetic stimulation of rostral DMHVgat neurons increased Cort, although with a lower magnitude compared to DMHVglut2 neuron stimulation, suggesting a role in disinhibiting PVHCRH neurons. Supporting this hypothesis, we found that rostral DMHVgat neurons project directly to GABAergic neurons in the caudal ventral part of the PVH and adjacent peri-PVH area (cvPVH), which directly inhibit PVHCRH neurons, and that activating the rostral DMHVgat terminals in the cvPVH in brain slices reduced GABAergic afferent input onto the PVHCRH neurons. Finally, ablation of cvPVHVgat neurons resulted in increased Cort release at the onset of the active phase, affirming the pivotal role of the DMHVgat → cvPVHVgat → PVHCRH pathway in Cort secretion. In summary, our study delineates two parallel pathways transmitting temporal information to PVHCRH neurons, collectively orchestrating the daily surge in Cort in anticipation of the active phase. These findings are crucial to understand the neural circuits regulating Cort secretion, shedding light on the mechanisms governing this physiological process and the coordinated interplay between the SCN, SPZ, DMH, and PVH.
Keywords: corticosterone; corticotropin-releasing hormone; dorsomedial hypothalamus and paraventricular hypothalamus.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures





References
-
- Focke C. M. B. & Iremonger K. J. Rhythmicity matters: Circadian and ultradian patterns of HPA axis activity. Mol. Cell. Endocrinol. 501, 110652 (2020). - PubMed
-
- Moore R. Y. & Eichler V. B. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 42, 201–206 (1972). - PubMed
-
- Buijs R. M., Markman M., Nunes-Cardoso B., Hou Y. X. & Shinn S. Projections of the suprachiasmatic nucleus to stress-related areas in the rat hypothalamus: a light and electron microscopic study. J. Comp. Neurol. 335, 42–54 (1993). - PubMed
-
- Dai J., Swaab D. F. & Buijs R. M. Distribution of vasopressin and vasoactive intestinal polypeptide (VIP) fibers in the human hypothalamus with special emphasis on suprachiasmatic nucleus efferent projections. J. Comp. Neurol. 383, 397–414 (1997). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
