

Dissemination of pathogenic bacteria is reinforced by a MARTX toxin effector duet
- PMID: 39043696
- PMCID: PMC11266601
- DOI: 10.1038/s41467-024-50650-0
Dissemination of pathogenic bacteria is reinforced by a MARTX toxin effector duet
Abstract
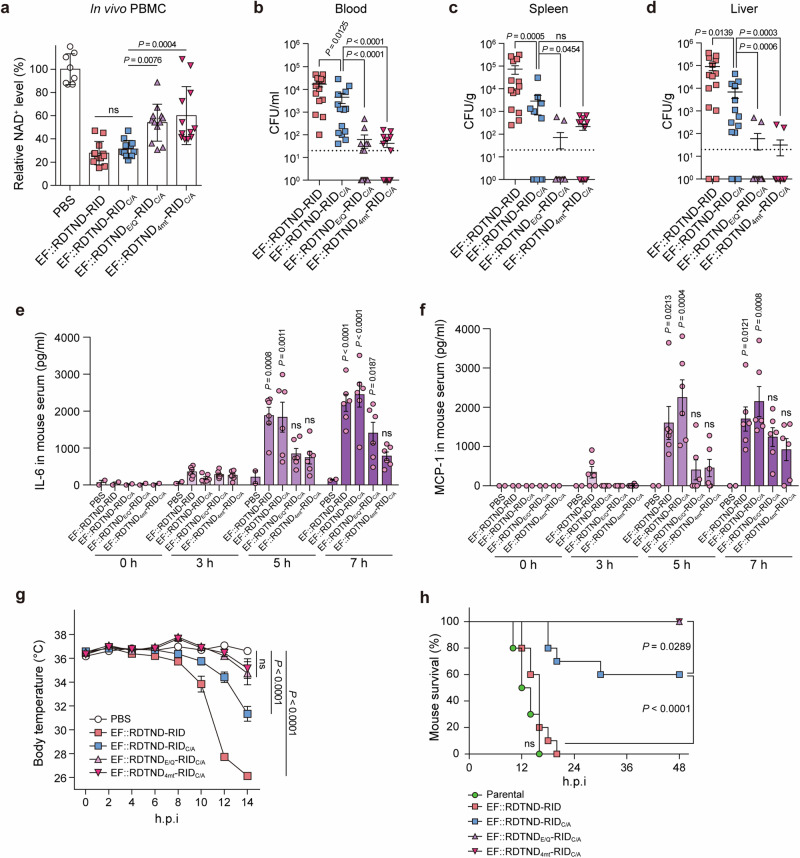
Multiple bacterial genera take advantage of the multifunctional autoprocessing repeats-in-toxin (MARTX) toxin to invade host cells. Secretion of the MARTX toxin by Vibrio vulnificus, a deadly opportunistic pathogen that causes primary septicemia, the precursor of sepsis, is a major driver of infection; however, the molecular mechanism via which the toxin contributes to septicemia remains unclear. Here, we report the crystal and cryo-electron microscopy (EM) structures of a toxin effector duet comprising the domain of unknown function in the first position (DUF1)/Rho inactivation domain (RID) complexed with human targets. These structures reveal how the duet is used by bacteria as a potent weapon. The data show that DUF1 acts as a RID-dependent transforming NADase domain (RDTND) that disrupts NAD+ homeostasis by hijacking calmodulin. The cryo-EM structure of the RDTND-RID duet complexed with calmodulin and Rac1, together with immunological analyses in vitro and in mice, provide mechanistic insight into how V. vulnificus uses the duet to suppress ROS generation by depleting NAD(P)+ and modifying Rac1 in a mutually-reinforcing manner that ultimately paralyzes first line immune responses, promotes dissemination of invaders, and induces sepsis. These data may allow development of tools or strategies to combat MARTX toxin-related human diseases.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
