

B cells secrete functional antigen-specific IgG antibodies on extracellular vesicles
- PMID: 39043800
- PMCID: PMC11266516
- DOI: 10.1038/s41598-024-67912-y
B cells secrete functional antigen-specific IgG antibodies on extracellular vesicles
Abstract
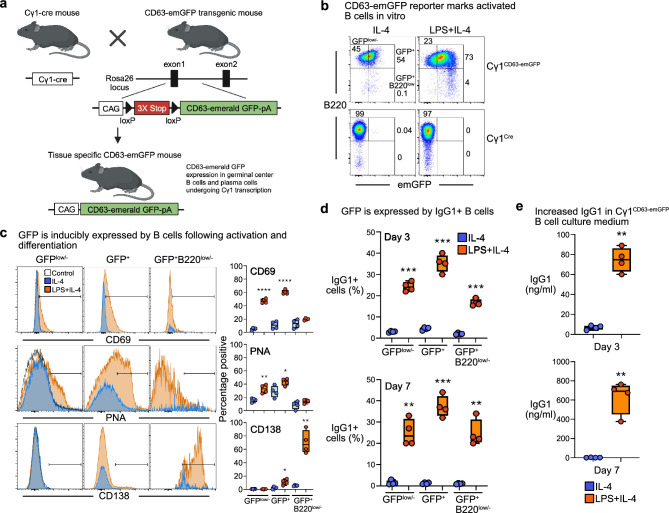
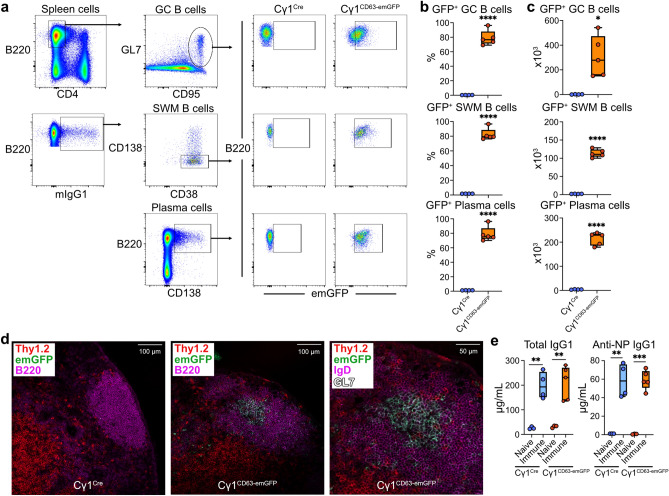
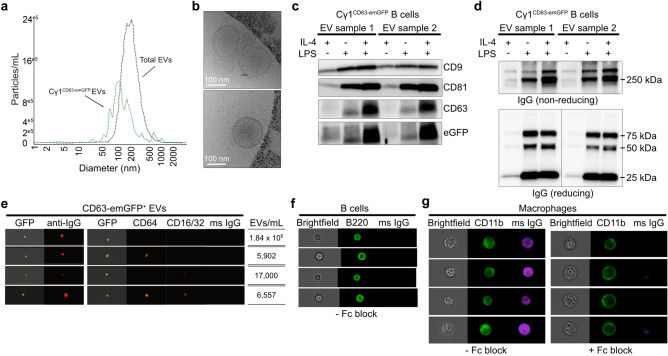
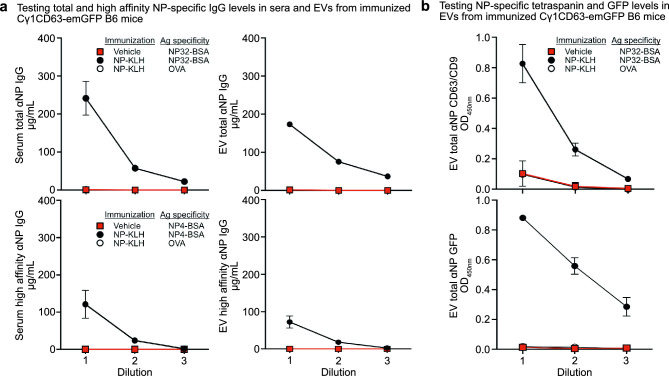
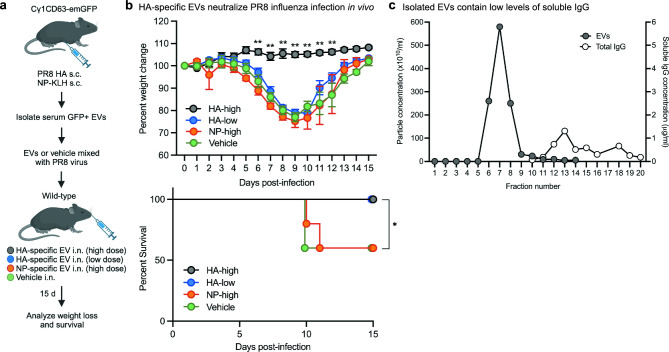
B cells and the antibodies they produce are critical in host defense against pathogens and contribute to various immune-mediated diseases. B cells responding to activating signals in vitro release extracellular vesicles (EV) that carry surface antibodies, yet B cell production of EVs that express antibodies and their function in vivo is incompletely understood. Using transgenic mice expressing the Cre recombinase in B cells switching to IgG1 to induce expression of fusion proteins between emerald green fluorescent protein (emGFP) and the EV tetraspanin CD63 as a model, we identify emGFP expression in B cells responding to foreign antigen in vivo and characterize the emGFP+ EVs they release. Our data suggests that emGFP+ germinal center B cells undergoing immunoglobulin class switching to express IgG and their progeny memory B cells and plasma cells, also emGFP+, are sources of circulating antigen-specific IgG+ EVs. Furthermore, using a mouse model of influenza virus infection, we find that IgG+ EVs specific for the influenza hemagglutinin antigen protect against virus infection. In addition, crossing the B cell Cre driver EV reporter mice onto the Nba2 lupus-prone strain revealed increased circulating emGFP+ EVs that expressed surface IgG against nuclear antigens linked to autoimmunity. These data identify EVs loaded with antibodies as a novel route for antibody secretion in B cells that contribute to adaptive immune responses, with important implications for different functions of IgG+ EVs in infection and autoimmunity.
Keywords: Adaptive immunity; Antibody production; B cells; Extracellular vesicles.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
