

Misaligned feeding uncouples daily rhythms within brown adipose tissue and between peripheral clocks
- PMID: 39046875
- PMCID: PMC12223410
- DOI: 10.1016/j.celrep.2024.114523
Misaligned feeding uncouples daily rhythms within brown adipose tissue and between peripheral clocks
Abstract
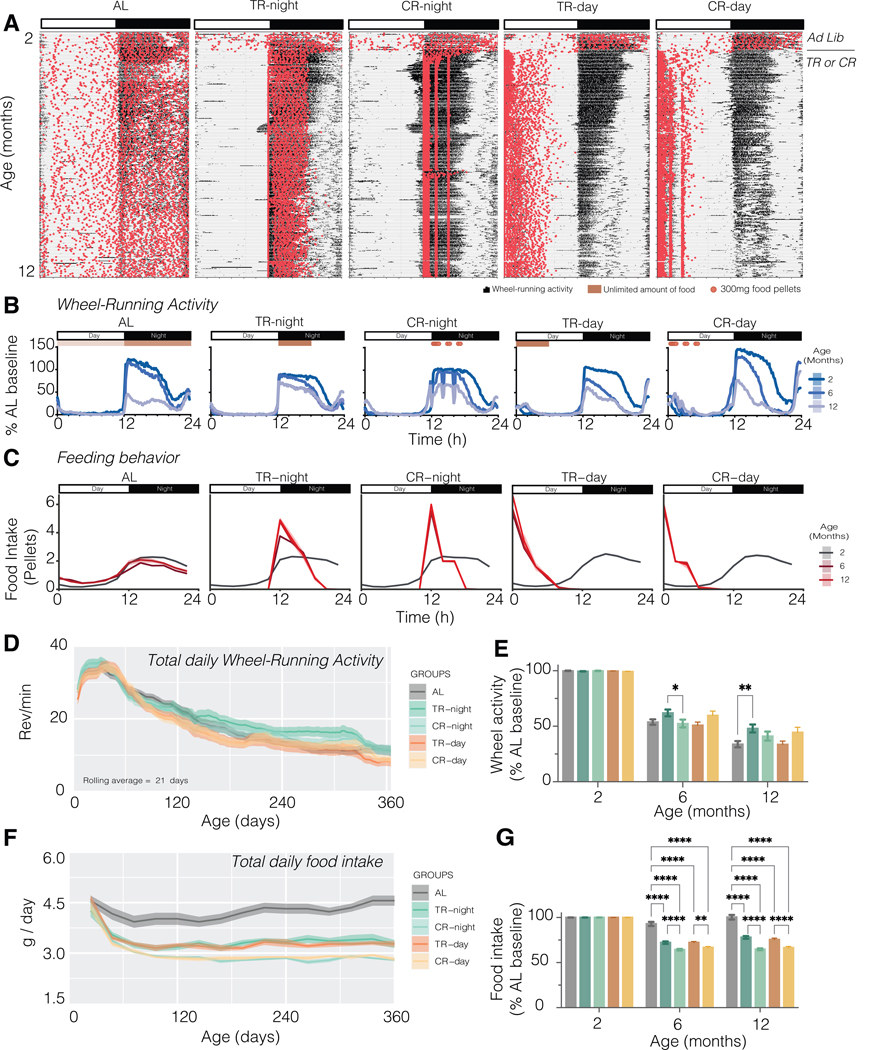
Extended food consumption during the rest period perturbs the phase relationship between circadian clocks in the periphery and the brain, leading to adverse health effects. Beyond the liver, how metabolic organs respond to a timed hypocaloric diet is largely unexplored. We investigated how feeding schedules impacted circadian gene expression in epididymal white and brown adipose tissue (eWAT and BAT) compared to the liver and hypothalamus. We restricted food to either daytime or nighttime in C57BL/6J male mice, with or without caloric restriction. Unlike the liver and eWAT, rhythmic clock genes in the BAT remained insensitive to feeding time, similar to the hypothalamus. We uncovered an internal split within the BAT in response to conflicting environmental cues, displaying inverted oscillations on a subset of metabolic genes without modifying its local core circadian machinery. Integrating tissue-specific responses on circadian transcriptional networks with metabolic outcomes may help elucidate the mechanism underlying the health burden of eating at unusual times.
Keywords: CP: Metabolism; brown adipose tissue; caloric restriction; circadian clocks; dietary interventions; liver; misaligned feeding; mouse behavior; time-restricted feeding.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
