

Machine Learning-Directed Conversion of Glioblastoma Cells to Dendritic Cell-Like Antigen-Presenting Cells as Cancer Immunotherapy
- PMID: 39051633
- PMCID: PMC11491168
- DOI: 10.1158/2326-6066.CIR-23-0721
Machine Learning-Directed Conversion of Glioblastoma Cells to Dendritic Cell-Like Antigen-Presenting Cells as Cancer Immunotherapy
Abstract
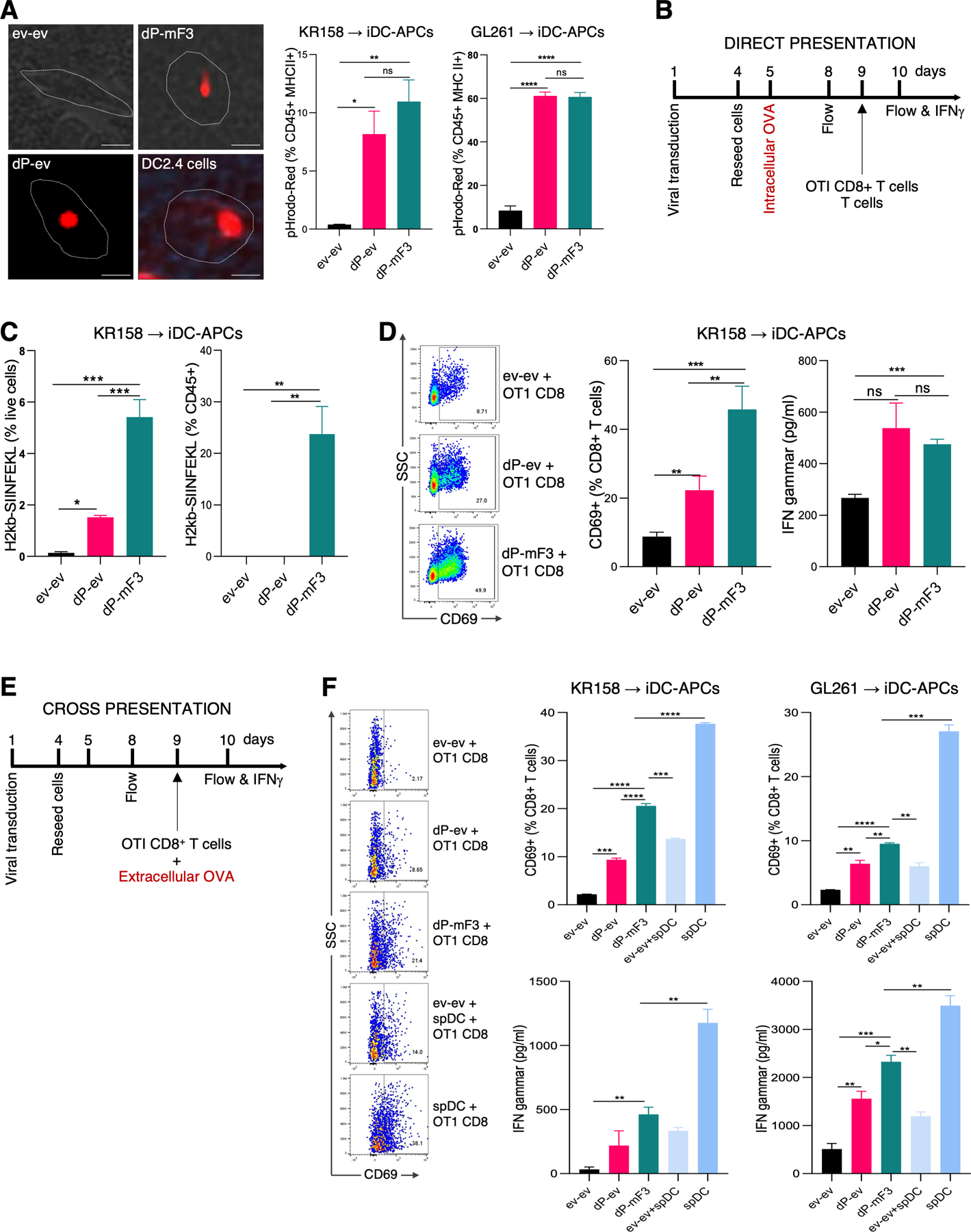
Immunotherapy has limited efficacy in glioblastoma (GBM) due to the blood-brain barrier and the immunosuppressed or "cold" tumor microenvironment (TME) of GBM, which is dominated by immune-inhibitory cells and depleted of CTL and dendritic cells (DC). Here, we report the development and application of a machine learning precision method to identify cell fate determinants (CFD) that specifically reprogram GBM cells into induced antigen-presenting cells with DC-like functions (iDC-APC). In murine GBM models, iDC-APCs acquired DC-like morphology, regulatory gene expression profile, and functions comparable to natural DCs. Among these acquired functions were phagocytosis, direct presentation of endogenous antigens, and cross-presentation of exogenous antigens. The latter endowed the iDC-APCs with the ability to prime naïve CD8+ CTLs, a hallmark DC function critical for antitumor immunity. Intratumor iDC-APCs reduced tumor growth and improved survival only in immunocompetent animals, which coincided with extensive infiltration of CD4+ T cells and activated CD8+ CTLs in the TME. The reactivated TME synergized with an intratumor soluble PD1 decoy immunotherapy and a DC-based GBM vaccine, resulting in robust killing of highly resistant GBM cells by tumor-specific CD8+ CTLs and significantly extended survival. Lastly, we defined a unique CFD combination specifically for the human GBM to iDC-APC conversion of both glioma stem-like cells and non-stem-like cell GBM cells, confirming the clinical utility of a computationally directed, tumor-specific conversion immunotherapy for GBM and potentially other solid tumors.
©2024 American Association for Cancer Research.
Conflict of interest statement
Figures






References
-
- Korman AJ, Garrett-Thomson SC, Lonberg N. The foundations of immune checkpoint blockade and the ipilimumab approval decennial. Nature Reviews Drug Discovery. 2022;21(7):509–28. - PubMed
-
- Medikonda R, Dunn G, Rahman M, Fecci P, Lim M. A review of glioblastoma immunotherapy. J Neurooncol 2021;151(1):41–53. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
