

Metastatic breast cancer cells are metabolically reprogrammed to maintain redox homeostasis during metastasis
- PMID: 39053265
- PMCID: PMC11321393
- DOI: 10.1016/j.redox.2024.103276
Metastatic breast cancer cells are metabolically reprogrammed to maintain redox homeostasis during metastasis
Abstract
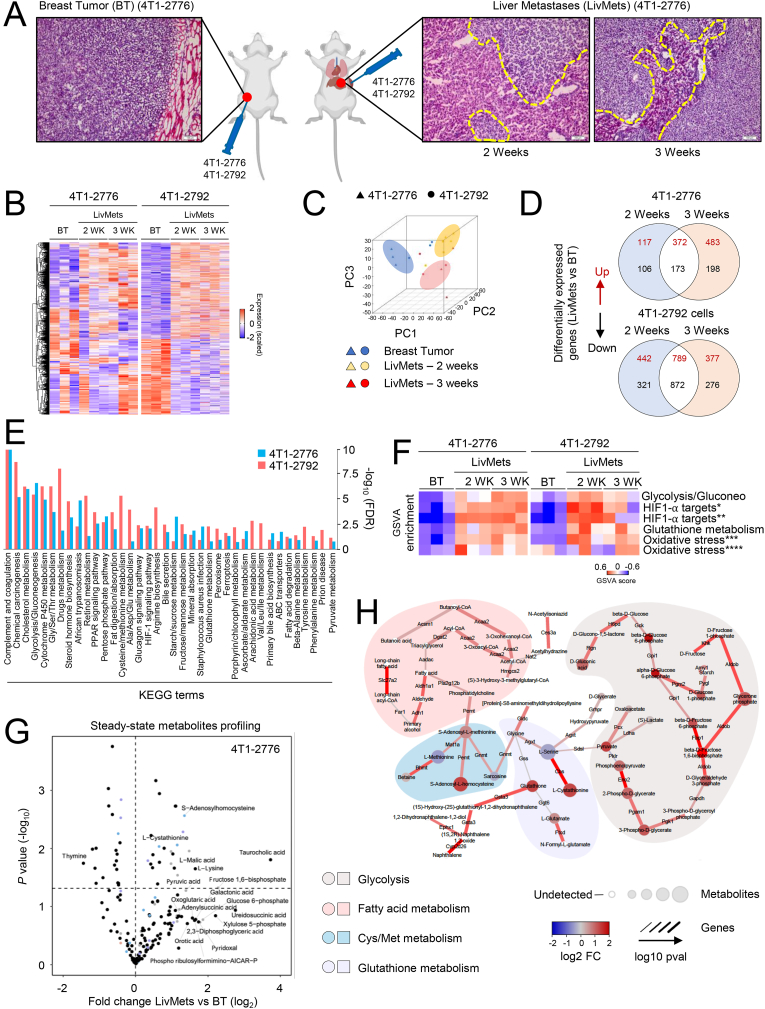
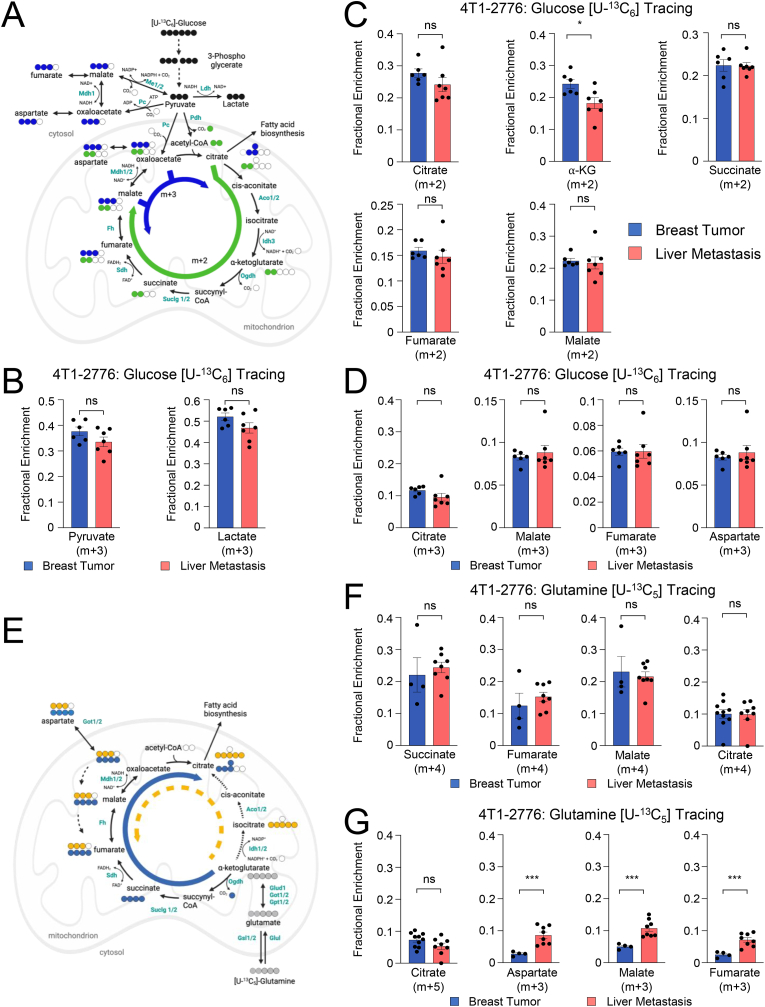
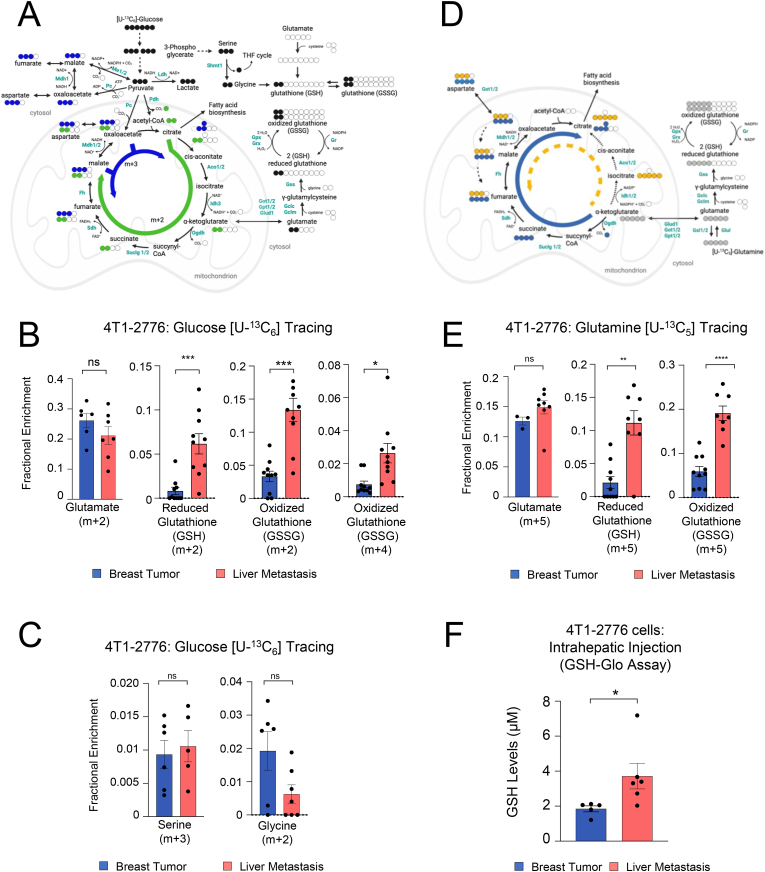
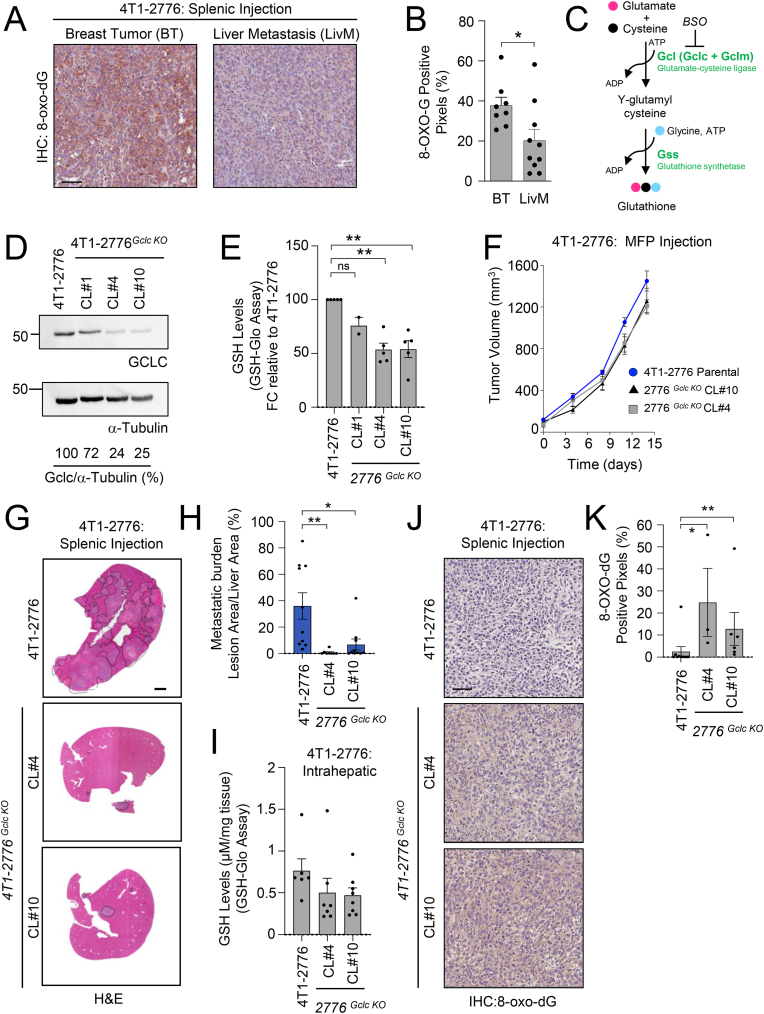
Metabolic rewiring is essential for tumor growth and progression to metastatic disease, yet little is known regarding how cancer cells modify their acquired metabolic programs in response to different metastatic microenvironments. We have previously shown that liver-metastatic breast cancer cells adopt an intrinsic metabolic program characterized by increased HIF-1α activity and dependence on glycolysis. Here, we confirm by in vivo stable isotope tracing analysis (SITA) that liver-metastatic breast cancer cells retain a glycolytic profile when grown as mammary tumors or liver metastases. However, hepatic metastases exhibit unique metabolic adaptations including elevated expression of genes involved in glutathione (GSH) biosynthesis and reactive oxygen species (ROS) detoxification when compared to mammary tumors. Accordingly, breast-cancer-liver-metastases exhibited enhanced de novo GSH synthesis. Confirming their increased capacity to mitigate ROS-mediated damage, liver metastases display reduced levels of 8-Oxo-2'-deoxyguanosine. Depletion of the catalytic subunit of the rate-limiting enzyme in glutathione biosynthesis, glutamate-cysteine ligase (GCLC), strongly reduced the capacity of breast cancer cells to form liver metastases, supporting the importance of these distinct metabolic adaptations. Loss of GCLC also affected the early steps of the metastatic cascade, leading to decreased numbers of circulating tumor cells (CTCs) and impaired metastasis to the liver and the lungs. Altogether, our results indicate that GSH metabolism could be targeted to prevent the dissemination of breast cancer cells.
Keywords: Breast cancer; GCLC; Glutathione; Glycolysis; HIF-1α; Liver metastasis; Metabolism; Oxidative stress.
Copyright © 2024. Published by Elsevier B.V.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
