

Altered neuronal lactate dehydrogenase A expression affects cognition in a sex- and age-dependent manner
- PMID: 39055955
- PMCID: PMC11269950
- DOI: 10.1016/j.isci.2024.110342
Altered neuronal lactate dehydrogenase A expression affects cognition in a sex- and age-dependent manner
Abstract
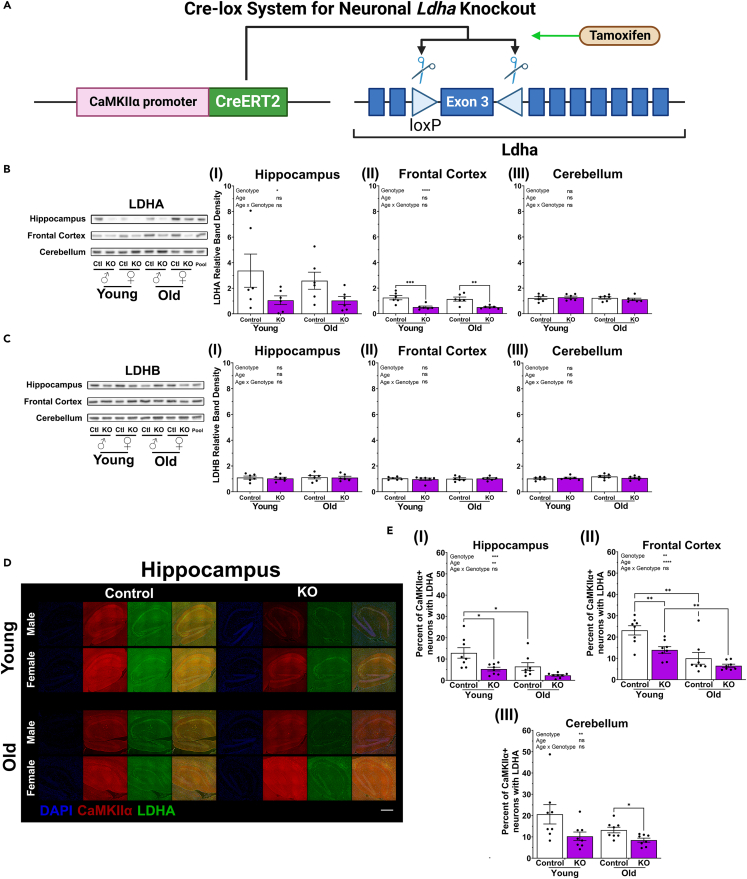
The astrocyte-neuron lactate shuttle (ANLS) model posits that astrocyte-generated lactate is transported to neurons to fuel memory processes. However, neurons express high levels of lactate dehydrogenase A (LDHA), the rate-limiting enzyme of lactate production, suggesting a cognitive role for neuronally generated lactate. It was hypothesized that lactate metabolism in neurons is critical for learning and memory. Here transgenic mice were generated to conditionally induce or knockout (KO) the Ldha gene in CNS neurons of adult mice. High pattern separation memory was enhanced by neuronal Ldha induction in young females, and by neuronal Ldha KO in aged females. In older mice, Ldha induction caused cognitive deficits whereas Ldha KO caused cognitive improvements. Genotype-associated cognitive changes were often only observed in one sex or oppositely in males and females. Thus, neuronal-generated lactate has sex-specific cognitive effects, is largely indispensable at young age, and may be detrimental to learning and memory with aging.
Keywords: Behavioral neuroscience; Cellular neuroscience; Molecular neuroscience; Neuroscience; Sensory neuroscience.
© 2024 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures









References
-
- Aiello L.C., Wheeler P. The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution. Curr. Anthropol. 1995;36:199–221. doi: 10.1086/204350. - DOI
-
- Clarke D.D., Sokoloff L. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 1999. Circulation and energy metabolism in the brain; pp. 637–669.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
