

Sterile production of interferons in the thymus affects T cell repertoire selection
- PMID: 39058762
- PMCID: PMC12052003
- DOI: 10.1126/sciimmunol.adp1139
Sterile production of interferons in the thymus affects T cell repertoire selection
Abstract
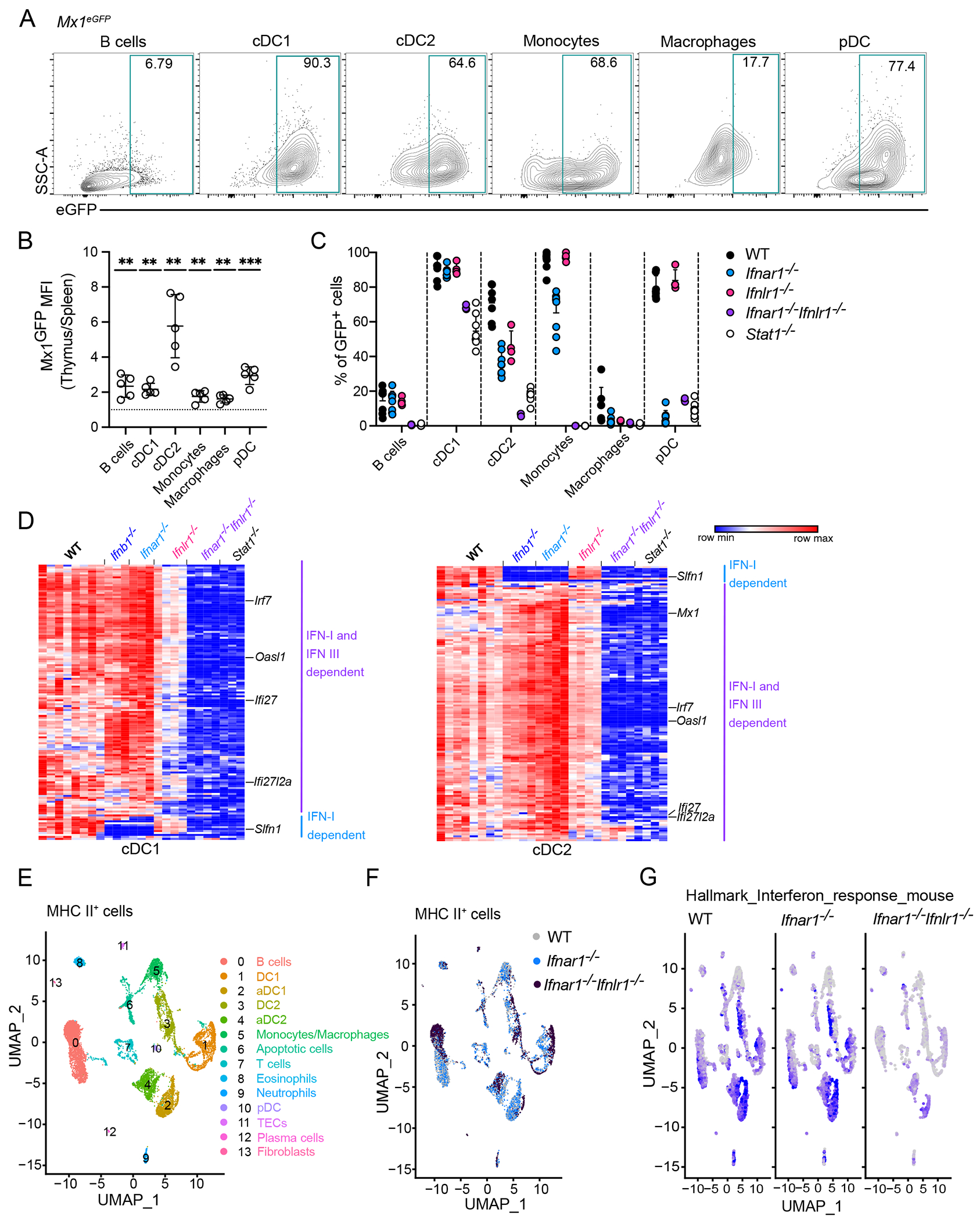
Type I and III interferons (IFNs) are robustly induced during infections and protect cells against viral infection. Both type I and III IFNs are also produced at low levels in the thymus at steady state; however, their role in T cell development and immune tolerance is unclear. Here, we found that both type I and III IFNs were constitutively produced by a very small number of AIRE+ murine thymic epithelial cells, independent of microbial stimulation. Antigen-presenting cells were highly responsive to thymic IFNs, and IFNs were required for the activation and maturation of thymic type 1 conventional dendritic cells, macrophages, and B cells. Loss of IFN sensing led to reduced regulatory T cell selection, reduced T cell receptor (TCR) repertoire diversity, and enhanced autoreactive T cell responses to self-antigens expressed during peripheral IFN signaling. Thus, constitutive exposure to IFNs in the thymus is required for generating a tolerant and diverse TCR repertoire.
Conflict of interest statement
Figures




References
-
- Benhammadi M, Mathé J, Dumont-Lagacé M, Kobayashi KS, Gaboury L, Brochu S, Perreault C, IFN-λ Enhances Constitutive Expression of MHC Class I Molecules on Thymic Epithelial Cells. J Immunol 205, 1268–1280 (2020). - PubMed
-
- Lienenklaus S, Cornitescu M, Zietara N, Łyszkiewicz M, Gekara N, Jabłónska J, Edenhofer F, Rajewsky K, Bruder D, Hafner M, Staeheli P, Weiss S, Novel reporter mouse reveals constitutive and inflammatory expression of IFN-beta in vivo. J Immunol 183, 3229–3236 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
