

Functional architecture of intracellular oscillations in hippocampal dendrites
- PMID: 39060234
- PMCID: PMC11282248
- DOI: 10.1038/s41467-024-50546-z
Functional architecture of intracellular oscillations in hippocampal dendrites
Abstract
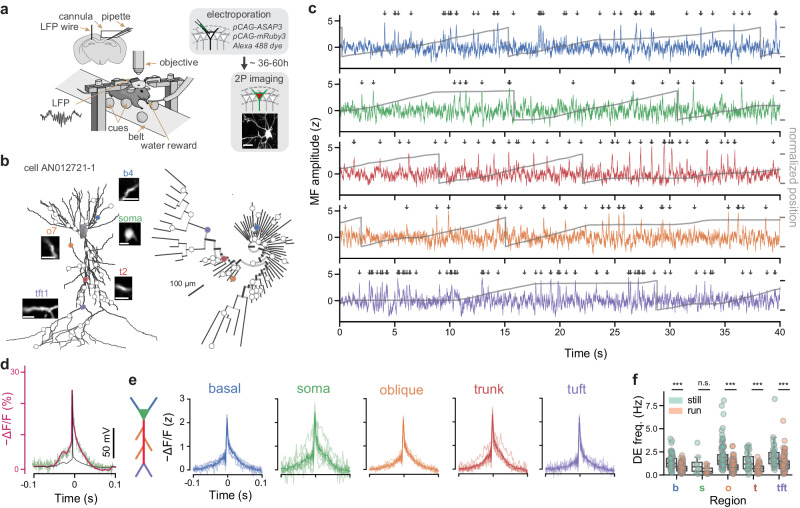
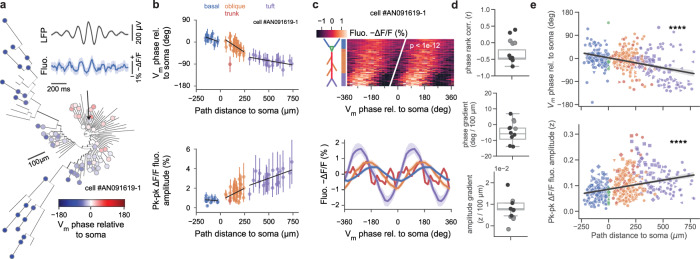
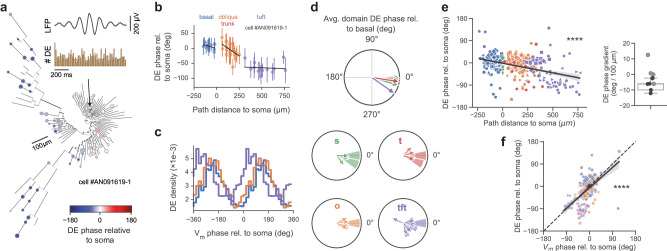
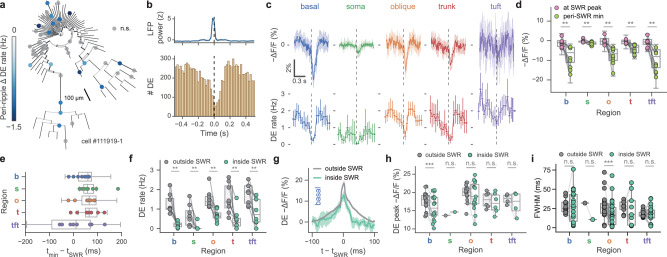
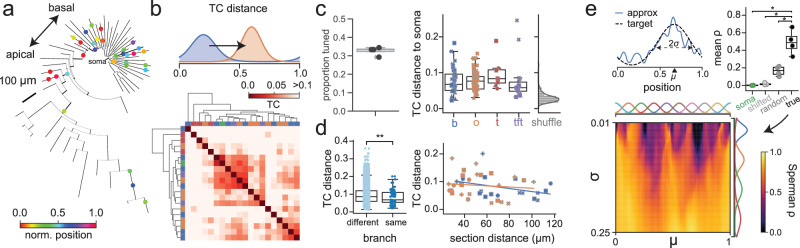
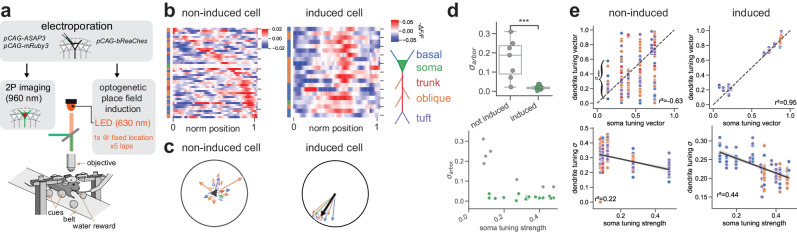
Fast electrical signaling in dendrites is central to neural computations that support adaptive behaviors. Conventional techniques lack temporal and spatial resolution and the ability to track underlying membrane potential dynamics present across the complex three-dimensional dendritic arbor in vivo. Here, we perform fast two-photon imaging of dendritic and somatic membrane potential dynamics in single pyramidal cells in the CA1 region of the mouse hippocampus during awake behavior. We study the dynamics of subthreshold membrane potential and suprathreshold dendritic events throughout the dendritic arbor in vivo by combining voltage imaging with simultaneous local field potential recording, post hoc morphological reconstruction, and a spatial navigation task. We systematically quantify the modulation of local event rates by locomotion in distinct dendritic regions, report an advancing gradient of dendritic theta phase along the basal-tuft axis, and describe a predominant hyperpolarization of the dendritic arbor during sharp-wave ripples. Finally, we find that spatial tuning of dendritic representations dynamically reorganizes following place field formation. Our data reveal how the organization of electrical signaling in dendrites maps onto the anatomy of the dendritic tree across behavior, oscillatory network, and functional cell states.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Functional architecture of intracellular oscillations in hippocampal dendrites.bioRxiv [Preprint]. 2024 Feb 12:2024.02.12.579750. doi: 10.1101/2024.02.12.579750. bioRxiv. 2024. Update in: Nat Commun. 2024 Jul 26;15(1):6295. doi: 10.1038/s41467-024-50546-z. PMID: 38405778 Free PMC article. Updated. Preprint.
References
MeSH terms
Grants and funding
- R01 MH124047/MH/NIMH NIH HHS/United States
- T32 NS064928/NS/NINDS NIH HHS/United States
- R01 MH124867/MH/NIMH NIH HHS/United States
- RF1 NS133381/NS/NINDS NIH HHS/United States
- T32 GM007367/GM/NIGMS NIH HHS/United States
- T32 MH020016/MH/NIMH NIH HHS/United States
- U01 NS103464/NS/NINDS NIH HHS/United States
- R01NS121106/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- U01NS115530/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- F99 NS139551/NS/NINDS NIH HHS/United States
- R01MH124047/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)
- F31 NS120783/NS/NINDS NIH HHS/United States
- R01 NS131728/NS/NINDS NIH HHS/United States
- R01MH124867/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)
- RF1 AG080818/AG/NIA NIH HHS/United States
- U01 NS115530/NS/NINDS NIH HHS/United States
- RF1 MH114105/MH/NIMH NIH HHS/United States
- R01 NS121106/NS/NINDS NIH HHS/United States
- R01NS133381/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
LinkOut - more resources
Full Text Sources
Miscellaneous
