

Bifidobacterium bifidum SAM-VI Riboswitch Conformation Change Requires Peripheral Helix Formation
- PMID: 39062457
- PMCID: PMC11274715
- DOI: 10.3390/biom14070742
Bifidobacterium bifidum SAM-VI Riboswitch Conformation Change Requires Peripheral Helix Formation
Abstract
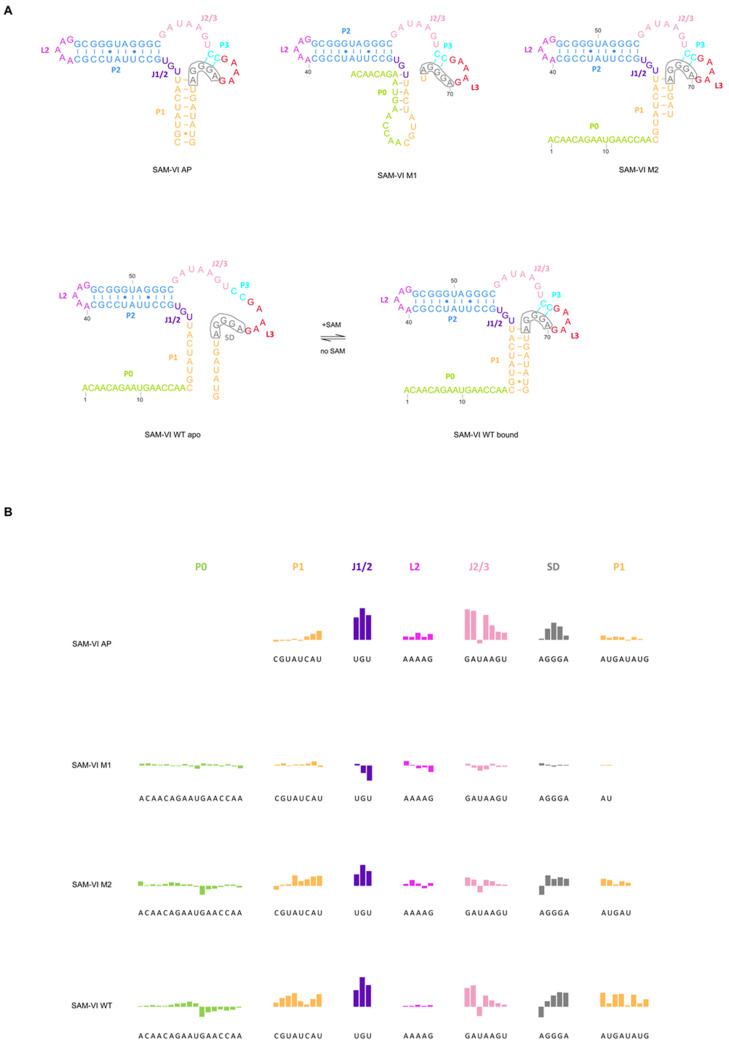
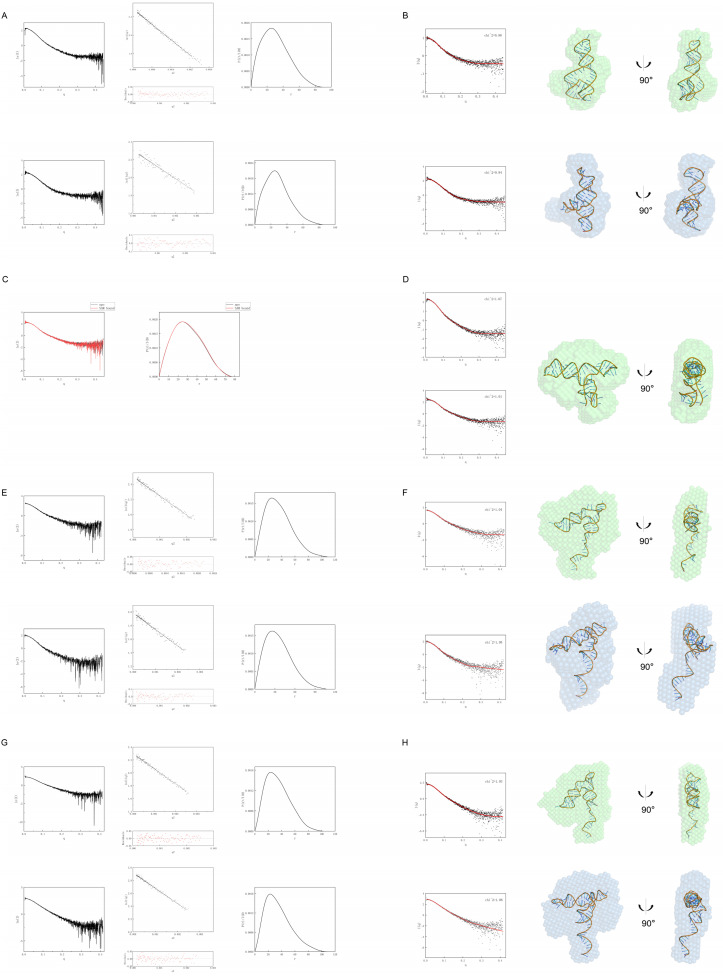
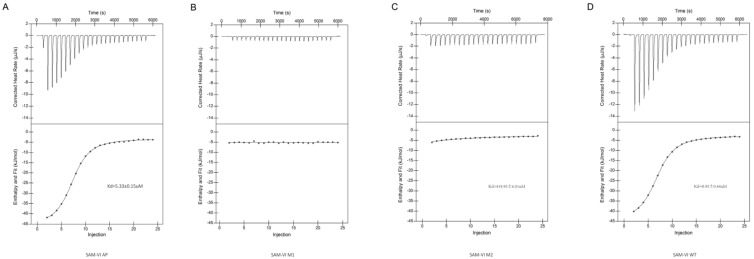
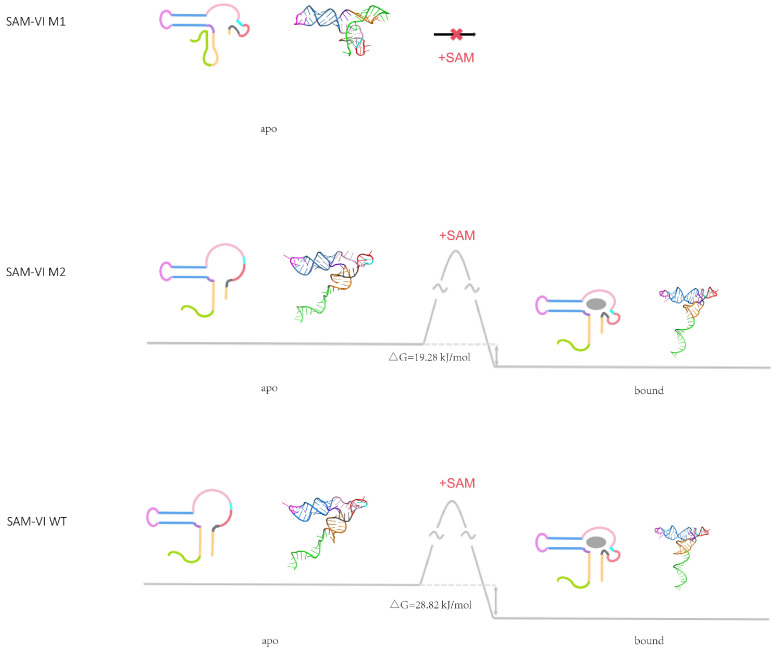
The Bifidobacterium bifidum SAM-VI riboswitch undergoes dynamic conformational changes that modulate downstream gene expression. Traditional structural methods such as crystallography capture the bound conformation at high resolution, and additional efforts would reveal details from the dynamic transition. Here, we revealed a transcription-dependent conformation model for Bifidobacterium bifidum SAM-VI riboswitch. In this study, we combine small-angle X-ray scattering, chemical probing, and isothermal titration calorimetry to unveil the ligand-binding properties and conformational changes of the Bifidobacterium bifidum SAM-VI riboswitch and its variants. Our results suggest that the SAM-VI riboswitch contains a pre-organized ligand-binding pocket and stabilizes into the bound conformation upon binding to SAM. Whether the P1 stem formed and variations in length critically influence the conformational dynamics of the SAM-VI riboswitch. Our study provides the basis for artificially engineering the riboswitch by manipulating its peripheral sequences without modifying the SAM-binding core.
Keywords: 3D modeling; S-adenosyl-methionine; SAM-VI riboswitch; SAXS; SHAPE; conformational dynamics; riboswitch.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
SAM-VI riboswitch structure and signature for ligand discrimination.Nat Commun. 2019 Dec 16;10(1):5728. doi: 10.1038/s41467-019-13600-9. Nat Commun. 2019. PMID: 31844059 Free PMC article.
-
Linker-Mediated Inactivation of the SAM-II Domain in the Tandem SAM-II/SAM-V Riboswitch.Int J Mol Sci. 2024 Oct 20;25(20):11288. doi: 10.3390/ijms252011288. Int J Mol Sci. 2024. PMID: 39457069 Free PMC article.
-
The impact of a ligand binding on strand migration in the SAM-I riboswitch.PLoS Comput Biol. 2013;9(5):e1003069. doi: 10.1371/journal.pcbi.1003069. Epub 2013 May 16. PLoS Comput Biol. 2013. PMID: 23704854 Free PMC article.
-
Structure-based insights into recognition and regulation of SAM-sensing riboswitches.Sci China Life Sci. 2023 Jan;66(1):31-50. doi: 10.1007/s11427-022-2188-7. Epub 2022 Nov 29. Sci China Life Sci. 2023. PMID: 36459353 Review.
-
Structural studies of the purine and SAM binding riboswitches.Cold Spring Harb Symp Quant Biol. 2006;71:259-68. doi: 10.1101/sqb.2006.71.015. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381305 Review.
Cited by
-
Observation of SAM-VI Riboswitch Dynamics Using Single-Molecule FRET.Biomolecules. 2025 Jun 9;15(6):841. doi: 10.3390/biom15060841. Biomolecules. 2025. PMID: 40563481 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
