

Enhanced ε-Poly-L-Lysine Production in Streptomyces albulus through Multi-Omics-Guided Metabolic Engineering
- PMID: 39062465
- PMCID: PMC11274744
- DOI: 10.3390/biom14070752
Enhanced ε-Poly-L-Lysine Production in Streptomyces albulus through Multi-Omics-Guided Metabolic Engineering
Abstract
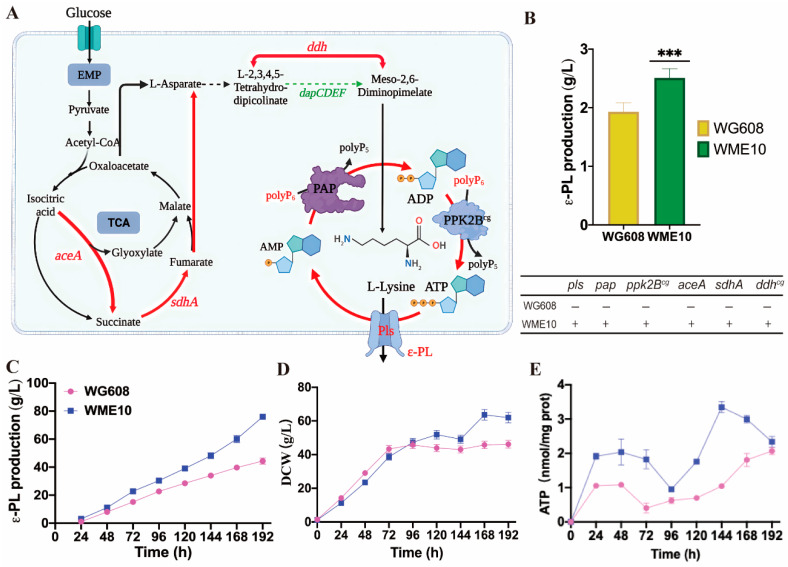
Safe and eco-friendly preservatives are crucial to preventing food spoilage and illnesses, as foodborne diseases caused by pathogens result in approximately 600 million cases of illness and 420,000 deaths annually. ε-Poly-L-lysine (ε-PL) is a novel food preservative widely used in many countries. However, its commercial application has been hindered by high costs and low production. In this study, ε-PL's biosynthetic capacity was enhanced in Streptomyces albulus WG608 through metabolic engineering guided by multi-omics techniques. Based on transcriptome and metabolome data, differentially expressed genes (fold change >2 or <0.5; p < 0.05) and differentially expressed metabolites (fold change >1.2 or <0.8) were separately subjected to gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. The integrative analysis of transcriptome, metabolome, and overexpression revealed the essential roles of isocitrate lyase, succinate dehydrogenase, flavoprotein subunit, diaminopimelate dehydrogenase, polyphosphate kinase, and polyP:AMP phosphotransferase in ε-PL biosynthesis. Subsequently, a strain with enhanced ATP supply, L-lysine supply, and ε-PL synthetase expression was constructed to improve its production. Finally, the resulting strain, S. albulus WME10, achieved an ε-PL production rate of 77.16 g/L in a 5 L bioreactor, which is the highest reported ε-PL production to date. These results suggest that the integrative analysis of the transcriptome and metabolome can facilitate the identification of key pathways and genetic elements affecting ε-PL synthesis, guiding further metabolic engineering and thus significantly enhancing ε-PL production. The method presented in this study could be applicable to other valuable natural antibacterial agents.
Keywords: antimicrobial; metabolic engineering; multi-omics; preservatives; ε-Poly-L-lysine.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





References
-
- Yeon C.S., Hye-Won N., Hanbyeol O., Mi K.Y., Seok S.W., Suncheol P., Wook-Jong J., Hongbaek C., Byung-Chul O., Jeongho P.J.N.A.R. Structural basis of flagellar motility regulation by the MogR repressor and the GmaR antirepressor in Listeria monocytogenes. Nucleic Acids Res. 2022;19:11315–11330. doi: 10.1093/nar/gkac815. - DOI - PMC - PubMed
-
- Bhattacharya S., Mishra S., Zuorro A., Salama H., Rusu A., Trif M. ε-Polylysine derived from marine bacteria-a possible natural preservative for raw milk storage. Fermentation. 2023;9:156. doi: 10.3390/fermentation9020156. - DOI
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
