

Guanidinium Chloride-Induced Haemolysis Assay to Measure New Permeation Pathway Functionality in Rodent Malaria Plasmodium berghei
- PMID: 39062495
- PMCID: PMC11274399
- DOI: 10.3390/biom14070781
Guanidinium Chloride-Induced Haemolysis Assay to Measure New Permeation Pathway Functionality in Rodent Malaria Plasmodium berghei
Abstract
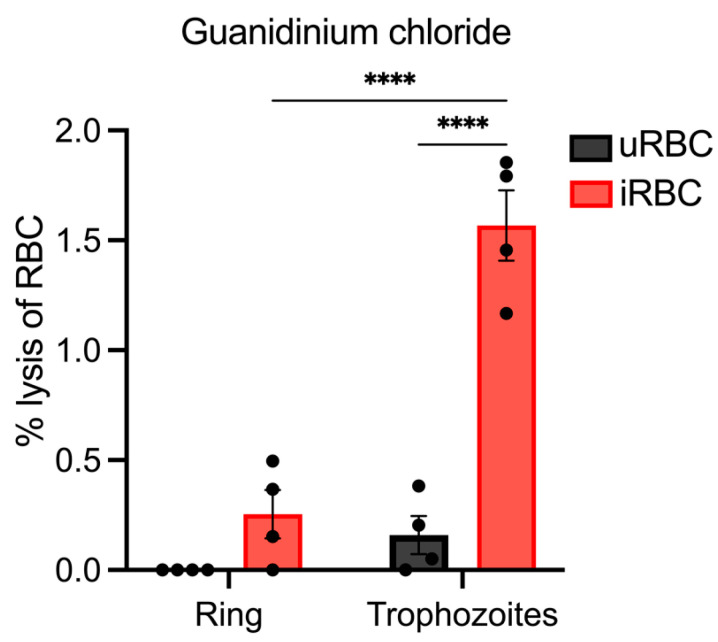
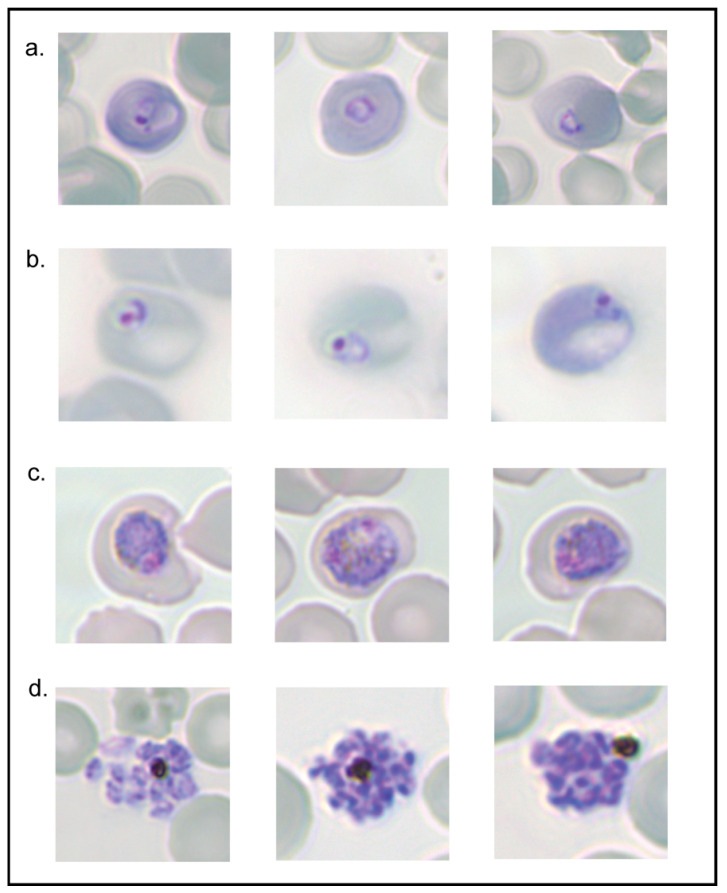
Parasite-derived new permeation pathways (NPPs) expressed at the red blood cell (RBC) membrane enable Plasmodium parasites to take up nutrients from the plasma to facilitate their survival. Thus, NPPs represent a potential novel therapeutic target for malaria. The putative channel component of the NPP in the human malaria parasite P. falciparum is encoded by mutually exclusively expressed clag3.1/3.2 genes. Complicating the study of the essentiality of these genes to the NPP is the addition of three clag paralogs whose contribution to the P. falciparum channel is uncertain. Rodent malaria P. berghei contains only two clag genes, and thus studies of P. berghei clag genes could significantly aid in dissecting their overall contribution to NPP activity. Previous methods for determining NPP activity in a rodent model have utilised flux-based assays of radioisotope-labelled substrates or patch clamping. This study aimed to ratify a streamlined haemolysis assay capable of assessing the functionality of P. berghei NPPs. Several isotonic lysis solutions were tested for their ability to preferentially lyse infected RBCs (iRBCs), leaving uninfected RBCs (uRBCs) intact. The osmotic lysis assay was optimised and validated in the presence of NPP inhibitors to demonstrate the uptake of the lysis solution via the NPPs. Guanidinium chloride proved to be the most efficient reagent to use in an osmotic lysis assay to establish NPP functionality. Furthermore, following treatment with guanidinium chloride, ring-stage parasites could develop into trophozoites and schizonts, potentially enabling use of guanidinium chloride for parasite synchronisation. This haemolysis assay will be useful for further investigation of NPPs in P. berghei and could assist in validating its protein constituents.
Keywords: Plasmodium berghei; Plasmodium surface anion channel; guanidinium chloride; new permeation pathway; osmotic lysis.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





Similar articles
-
Utilisation of an in vivo malaria model to provide functional proof for RhopH1/CLAG essentiality and conserved orthology with P. falciparum.J Biomed Sci. 2025 Feb 3;32(1):13. doi: 10.1186/s12929-024-01105-7. J Biomed Sci. 2025. PMID: 39894870 Free PMC article.
-
Proteomic and genetic analyses demonstrate that Plasmodium berghei blood stages export a large and diverse repertoire of proteins.Mol Cell Proteomics. 2013 Feb;12(2):426-48. doi: 10.1074/mcp.M112.021238. Epub 2012 Nov 28. Mol Cell Proteomics. 2013. PMID: 23197789 Free PMC article.
-
Plasmodium berghei K13 Mutations Mediate In Vivo Artemisinin Resistance That Is Reversed by Proteasome Inhibition.mBio. 2020 Nov 10;11(6):e02312-20. doi: 10.1128/mBio.02312-20. mBio. 2020. PMID: 33173001 Free PMC article.
-
Sequestration and tissue accumulation of human malaria parasites: can we learn anything from rodent models of malaria?PLoS Pathog. 2010 Sep 30;6(9):e1001032. doi: 10.1371/journal.ppat.1001032. PLoS Pathog. 2010. PMID: 20941396 Free PMC article. Review.
-
Epigenetics of malaria parasite nutrient uptake, but why?Trends Parasitol. 2022 Aug;38(8):618-628. doi: 10.1016/j.pt.2022.05.005. Epub 2022 May 28. Trends Parasitol. 2022. PMID: 35641406 Free PMC article. Review.
Cited by
-
Utilisation of an in vivo malaria model to provide functional proof for RhopH1/CLAG essentiality and conserved orthology with P. falciparum.J Biomed Sci. 2025 Feb 3;32(1):13. doi: 10.1186/s12929-024-01105-7. J Biomed Sci. 2025. PMID: 39894870 Free PMC article.
References
-
- Straimer J., Gnädig N.F., Witkowski B., Amaratunga C., Duru V., Ramadani A.P., Dacheux M., Khim N., Zhang L., Lam S., et al. K13-propeller mutations confer artemisinin resistance in Plasmodium falciparum clinical isolates. Science. 2015;347:428–431. doi: 10.1126/science.1260867. - DOI - PMC - PubMed
-
- Uwimana A., Legrand E., Stokes B.H., Ndikumana J.-L.M., Warsame M., Umulisa N., Ngamije D., Munyaneza T., Mazarati J.-B., Munguti K., et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat. Med. 2020;26:1602–1608. doi: 10.1038/s41591-020-1005-2. - DOI - PMC - PubMed
-
- Uwimana A., Umulisa N., Venkatesan M., Svigel S.S., Zhou Z., Munyaneza T., Habimana R.M., Rucogoza A., Moriarty L.F., Sandford R., et al. Association of Plasmodium falciparum kelch13 R561H genotypes with delayed parasite clearance in Rwanda: An open-label, single-arm, multicentre, therapeutic efficacy study. Lancet Infect. Dis. 2021;21:1120–1128. doi: 10.1016/S1473-3099(21)00142-0. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
