

Morphological and Functional Alterations in the CA1 Pyramidal Neurons of the Rat Hippocampus in the Chronic Phase of the Lithium-Pilocarpine Model of Epilepsy
- PMID: 39062811
- PMCID: PMC11276980
- DOI: 10.3390/ijms25147568
Morphological and Functional Alterations in the CA1 Pyramidal Neurons of the Rat Hippocampus in the Chronic Phase of the Lithium-Pilocarpine Model of Epilepsy
Abstract
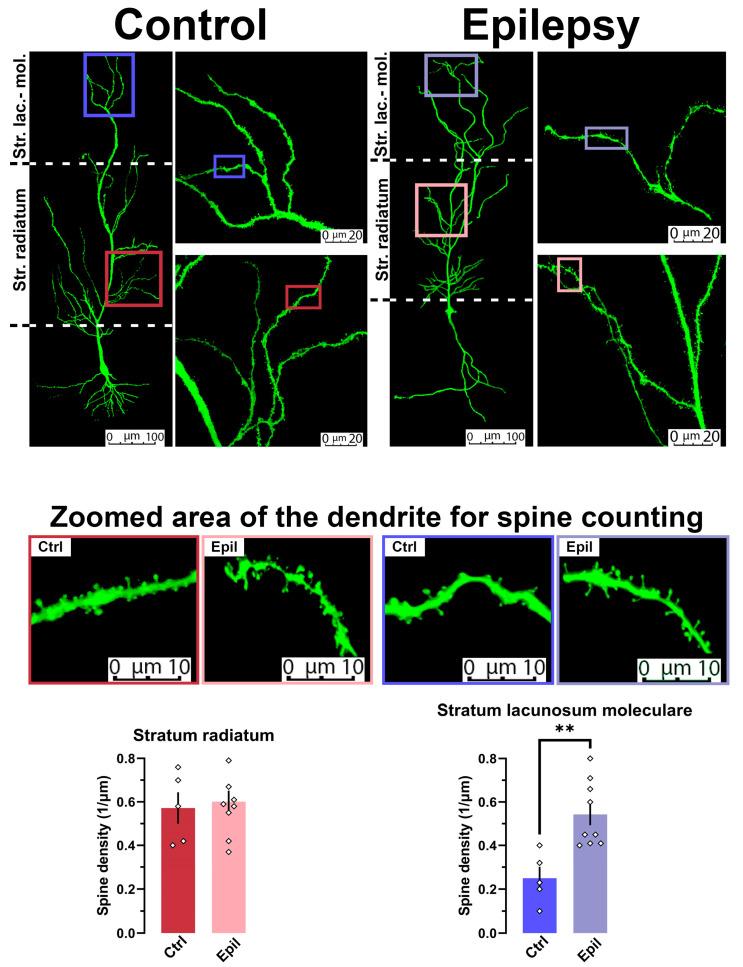
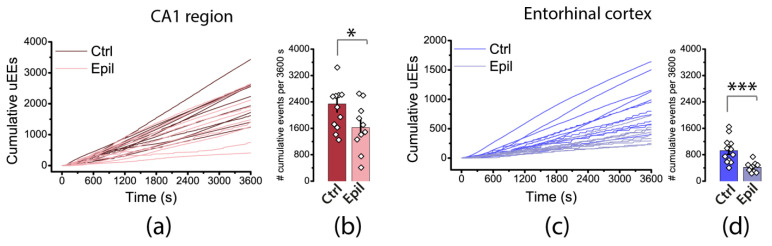
Epilepsy is known to cause alterations in neural networks. However, many details of these changes remain poorly understood. The objective of this study was to investigate changes in the properties of hippocampal CA1 pyramidal neurons and their synaptic inputs in a rat lithium-pilocarpine model of epilepsy. In the chronic phase of the model, we found a marked loss of pyramidal neurons in the CA1 area. However, the membrane properties of the neurons remained essentially unaltered. The results of the electrophysiological and morphological studies indicate that the direct pathway from the entorhinal cortex to CA1 neurons is reinforced in epileptic animals, whereas the inputs to them from CA3 are either unaltered or even diminished. In particular, the dendritic spine density in the str. lacunosum moleculare, where the direct pathway from the entorhinal cortex terminates, was found to be 2.5 times higher in epileptic rats than in control rats. Furthermore, the summation of responses upon stimulation of the temporoammonic pathway was enhanced by approximately twofold in epileptic rats. This enhancement is believed to be a significant contributing factor to the heightened epileptic activity observed in the entorhinal cortex of epileptic rats using an ex vivo 4-aminopyridine model.
Keywords: 4-aminopyridine model; action potential; entorhinal cortex; hippocampus; ictal discharge; pyramidal neuron; temporal lobe epilepsy.
Conflict of interest statement
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures








References
-
- World Health Organization Epilepsy. [(accessed on 29 January 2024)]. Available online: https://www.who.int/news-room/fact-sheets/detail/epilepsy.
-
- Wong R.K., Traub R.D., Miles R. Cellular Basis of Neuronal Synchrony in Epilepsy. Adv. Neurol. 1986;44:583–592. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
