

Movement-related increases in subthalamic activity optimize locomotion
- PMID: 39068661
- PMCID: PMC11407793
- DOI: 10.1016/j.celrep.2024.114495
Movement-related increases in subthalamic activity optimize locomotion
Abstract
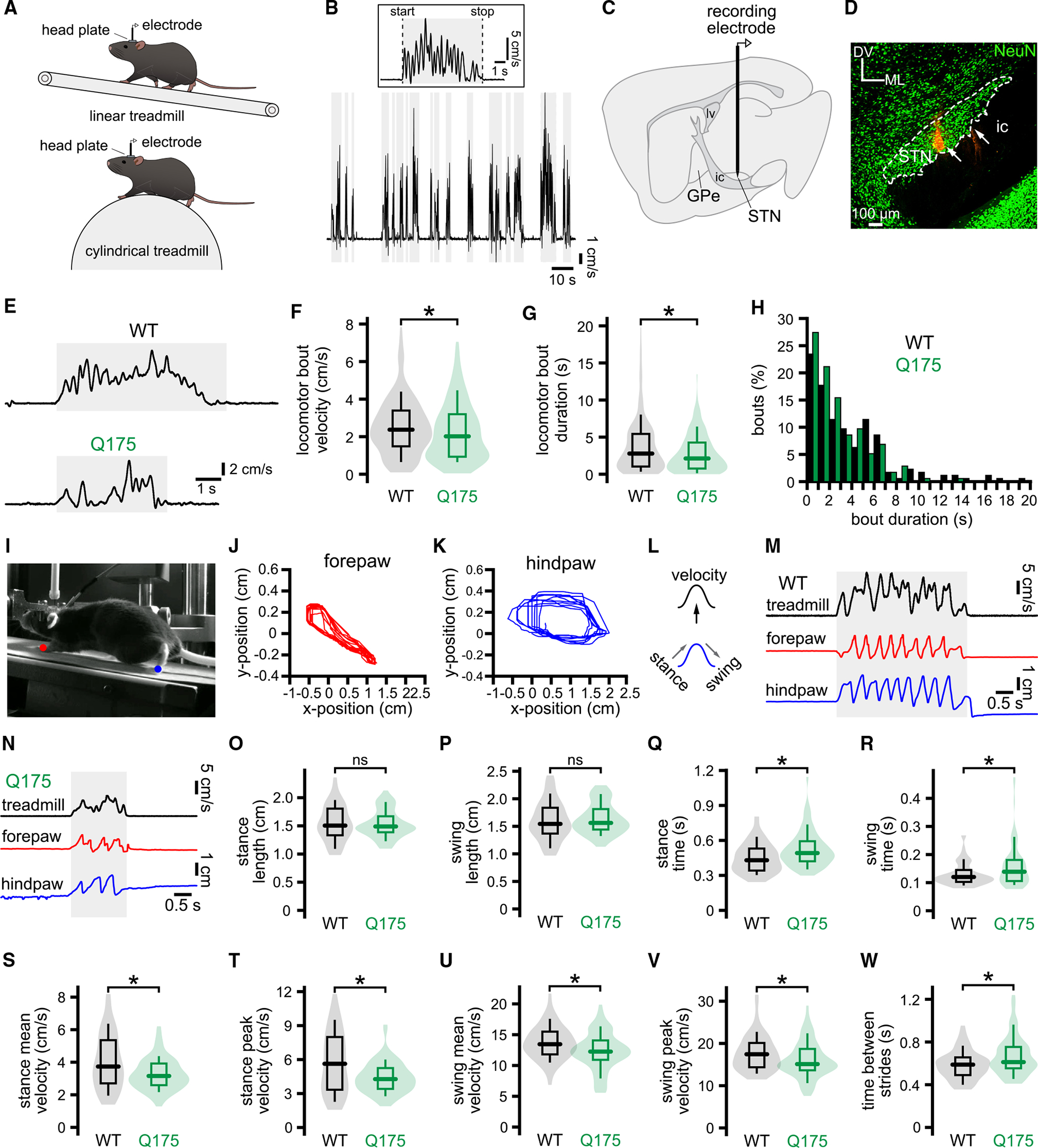
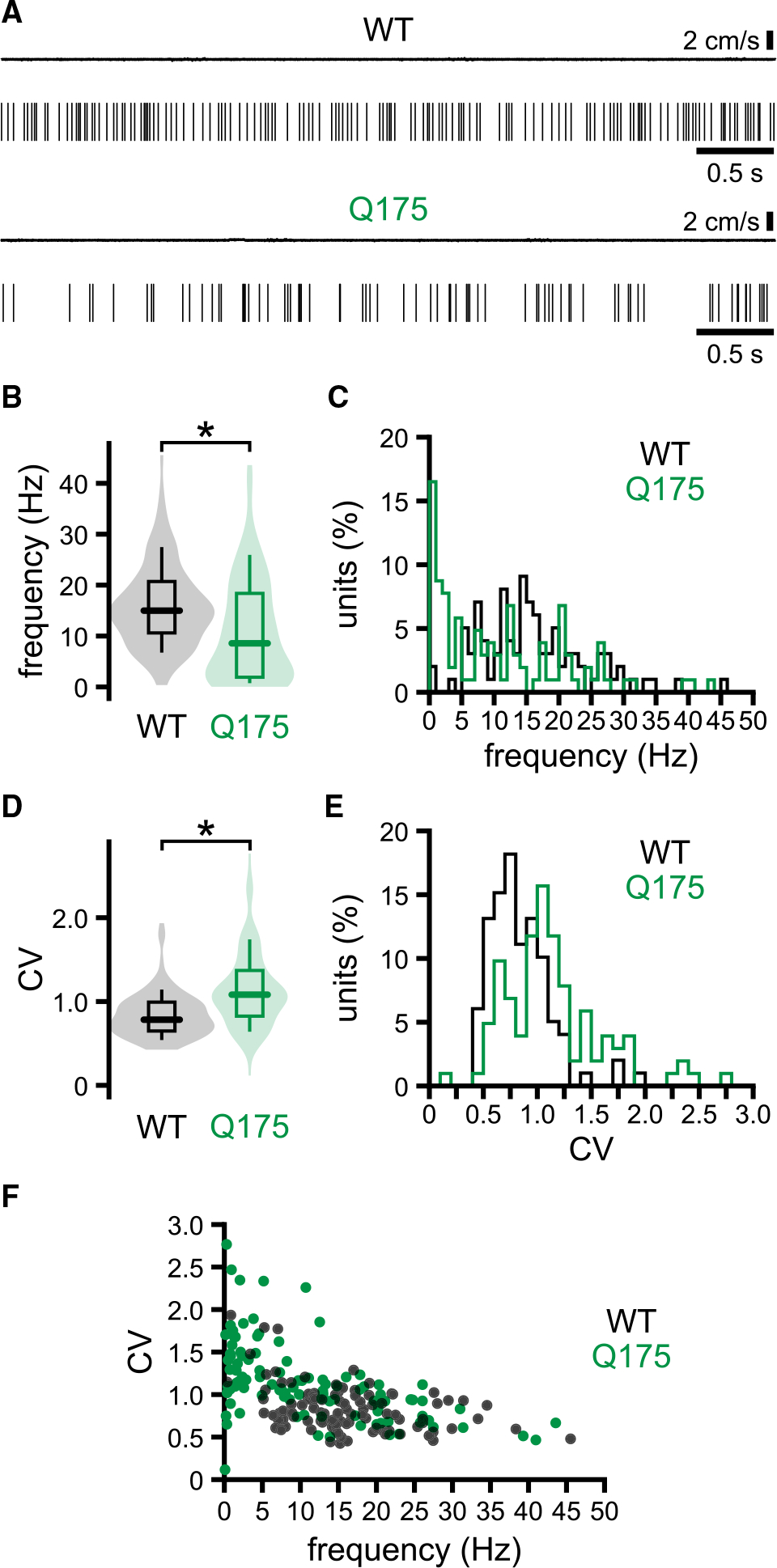
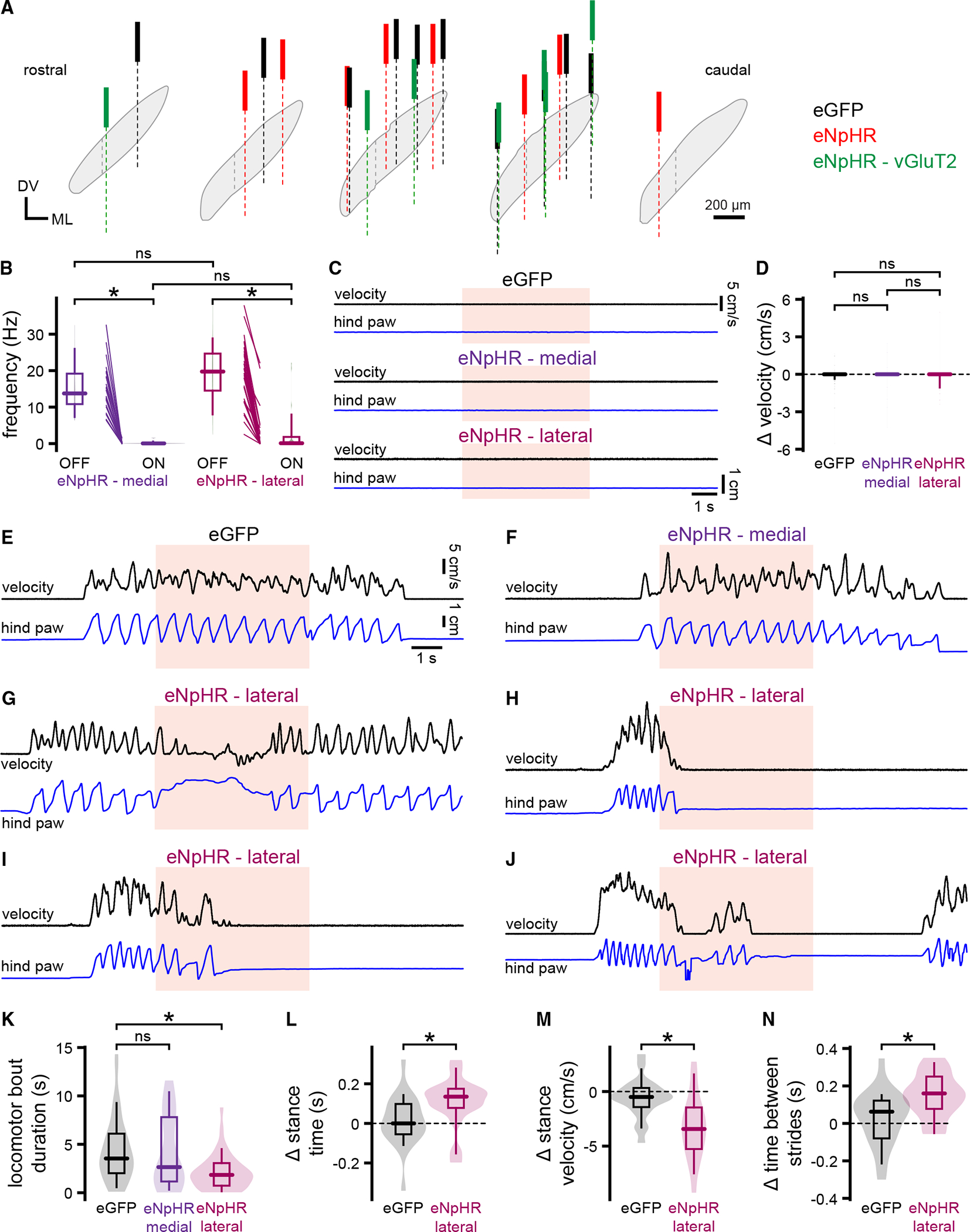
The subthalamic nucleus (STN) is traditionally thought to restrict movement. Lesion or prolonged STN inhibition increases movement vigor and propensity, while optogenetic excitation has opposing effects. However, STN neurons often exhibit movement-related increases in firing. To address this paradox, STN activity was recorded and manipulated in head-fixed mice at rest and during self-initiated and self-paced treadmill locomotion. We found that (1) most STN neurons (type 1) exhibit locomotion-dependent increases in activity, with half firing preferentially during the propulsive phase of the contralateral locomotor cycle; (2) a minority of STN neurons exhibit dips in activity or are uncorrelated with movement; (3) brief optogenetic inhibition of the lateral STN (where type 1 neurons are concentrated) slows and prematurely terminates locomotion; and (4) in Q175 Huntington's disease mice, abnormally brief, low-velocity locomotion is associated with type 1 hypoactivity. Together, these data argue that movement-related increases in STN activity contribute to optimal locomotor performance.
Keywords: CP: Neuroscience; Huntington’s disease; Parkinson’s disease; action execution; basal ganglia; deep brain stimulation; gait; hyperdirect pathway; indirect pathway; motor control; neuromodulation.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




Update of
-
Movement-related increases in subthalamic activity optimize locomotion.bioRxiv [Preprint]. 2024 May 28:2023.12.07.570617. doi: 10.1101/2023.12.07.570617. bioRxiv. 2024. Update in: Cell Rep. 2024 Aug 27;43(8):114495. doi: 10.1016/j.celrep.2024.114495. PMID: 38105984 Free PMC article. Updated. Preprint.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
