

Engineering single-cycle MeV vector for CRISPR-Cas9 gene editing
- PMID: 39070290
- PMCID: PMC11283025
- DOI: 10.1016/j.omtm.2024.101290
Engineering single-cycle MeV vector for CRISPR-Cas9 gene editing
Abstract
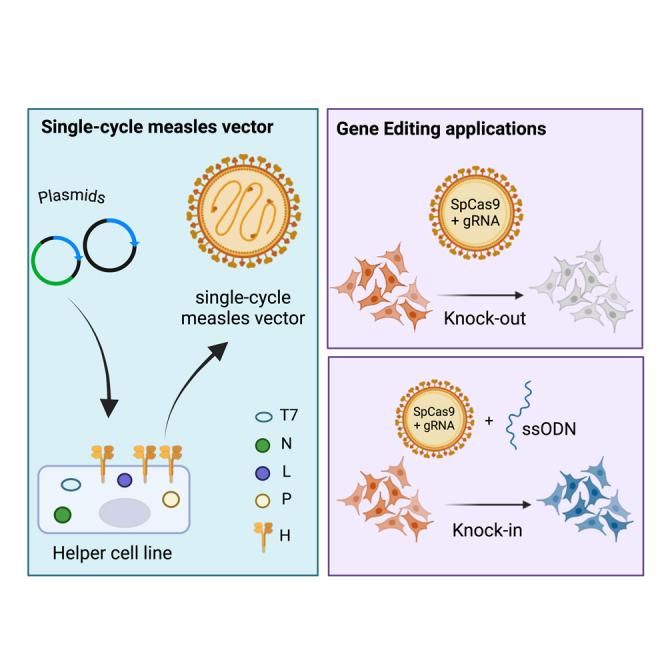
CRISPR-Cas9-mediated gene editing has vast applications in basic and clinical research and is a promising tool for several disorders. Our lab previously developed a non-integrating RNA virus, measles virus (MeV), as a single-cycle reprogramming vector by replacing the viral attachment protein with the reprogramming factors for induced pluripotent stem cell generation. Encouraged by the MeV reprogramming vector efficiency, in this study, we develop a single-cycle MeV vector to deliver the gRNA(s) and Cas9 nuclease to human cells for efficient gene editing. We show that the MeV vector achieved on-target gene editing of the reporter (mCherry) and endogenous genes (HBB and FANCD1) in human cells. Additionally, the MeV vector achieved precise knock-in via homology-directed repair using a single-stranded oligonucleotide donor. The MeV vector is a new and flexible platform for gene knock-out and knock-in modifications in human cells, capable of incorporating new technologies as they are developed.
Keywords: CRISPR; Cas9; HDR; NHEJ; gene editing; iPSC; measles; viral vector.
© 2024 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Ishino Y., Shinagawa H., Makino K., Amemura M., Nakata A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987;169:5429–5433. doi: 10.1128/jb.169.12.5429-5433.1987. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
